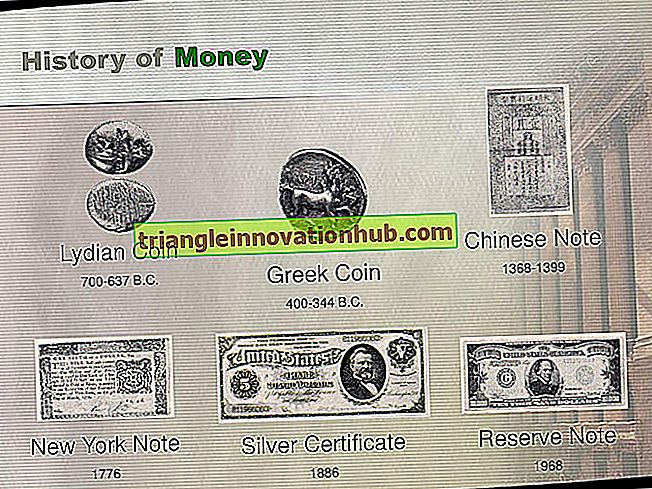
Fortpflanzungssystem bei Fischen
In diesem Artikel werden wir über die Fortpflanzungssysteme bei Fischen diskutieren.
Die Fortpflanzung bei Fischen ist bisexuell, hermaphroditisch oder parthenogen. Der vorherrschende Prozess ist die bisexuelle Wiedergabe. Bei solchen Fischen sind die Geschlechter getrennt, zB sind die Individuen zweihäusig. Bei einigen Fischen sind beide Geschlechter im gleichen Individuum vorhanden, dh solche Fische sind Hermaphrodite.
Die Beispiele sind Perca, Stizostedion, Micropterus. Bei einigen Fischen wurde ein jugendlicher Hermaphroditismus beobachtet. Bei der Poe-cilia Formosa-Parthenogenese tritt die Gynogenese auf, dh die Entwicklung von Jungen ohne Befruchtung.
Die Gonaden von Elasmobranchen entwickelten sich durch die Migration von primordialen Keimzellen aus dem extraembryonalen Endoderm, die aus dem Peritonealepithel und dem inter-renalen Analogon stammen. In Teleost stammen auch die Urkeimzellen vom Genitalrücken (als Schwellung). Die Urkeimzelle ist ziemlich groß und hat eine leicht ovale Form mit einem großen kugelförmigen Kern, der einen großen Kern enthält.
Der Genitalrücken entwickelt sich bei Wirbeltieren zu Kortex und Medulla. Gonaden in Teleostern und Cyclostomen bestehen jedoch nur aus Kortex und haben kein Markgewebe. In elasmobranch entwickelt sie sich zu Medulla, bei Frauen aber zur Kortikalis. Später durchlaufen die primordialen Keimzellen eine Meiose und vermehren sich und differenzieren sich schließlich in männliche und weibliche Gonaden.
Die Fische haben ein Paar bilateraler Gonaden. Sie sind im Allgemeinen symmetrisch. Sie werden von Mesenteries über dem Dach der Körperhöhle in enger Verbindung mit den Nieren am dorsalen Teil der Körperhöhle aufgehängt. Das Mesenterium bei Frauen heißt Mesovarium. Sie sind auch in Neoceratodus vorhanden (Abb. 20.1a, b, c). Während des Mannes ist es als Mesorchium bekannt. Es ist reich an Blutgefäßen und Nervenfasern.

Hämatopoetisches Epigonalorgel:
In allen Elasmobranchen sind Gonaden mit hämatopoetischen Epigonalorganen assoziiert. Die Hoden wie die Eierstöcke sind auch mit epigonalen Organen assoziiert (Abb. 20.1b, c). Sie reichen zurück bis zur Kloake, sie kann bei Heptanchus relativ klein sein oder bei Squalus fehlen. In Scyllium sind die Hoden mit epigonalen Organen verschmolzen.
Es ist in Cetorhinus gepaart, der rechts von hinten liegende und leicht dorsal am Eierstock und durch eine hintere Erweiterung des Mesovariums aufgehängt. Es ist etwa 60 cm lang und hat einen Durchmesser von 20 cm. Der vordere Bereich ist eng mit dem hinteren Drittel des Ovars verschmolzen.
Das linke epigonale Organ der Frau ist in Größe und Form ähnlich, obwohl es nicht mit einer Gonade assoziiert ist. Bei weiblichen Haien vieler Gattungen entwickelt sich nur ein Eierstock (Scyliorhinus, Pristiurus, Galeus, Mustelus), und daher ist das weibliche Eipgonalorgan in Übereinstimmung damit oft ungepaart, so dass sie auf der einen Seite besser sind als auf der anderen.
Weibliches Fortpflanzungsorgan:
Die weiblichen Fortpflanzungsorgane bestehen aus den Eierstöcken, den Eileitern und in einigen Fischen pseudo-kopulierenden Papillen (Abb. 20.2 und Abb. 20.3). Der Eierstock in Bezug auf Eileiter und Transport von Eizellen wird in zytovariater Typ, semicytovarian Typ und gymnovarian Typ unterschieden. Beim Zytovariern wird das Lumen des Eierstocks mit dem Eileiter verbunden und durch Genitalporen im Wasser freigesetzt.


Beim semicytovarischen Typ gehen die Eizellen anstelle des Eileiters durch eine trichterförmige transparente Rille, die sich in die Genitalpore öffnet. Ein solcher Zustand ist in Notopteridae, Osteoglassidae usw. vorhanden, wo der Eileiter teilweise oder vollständig degeneriert und daher Eizellen in die Coelomhöhle geschüttet und dann durch Poren oder Trichter befördert werden.
Der gymnovarische Ovarialtyp ist nicht durchgehend mit dem Eileiter verbunden. Der Eierstock hängt wie ein Vorhang herab und die Oozyten werden in die Coelom- oder Peritonealhöhle ovuliert und durch Zilien zum Ovidukt gebracht.
Normalerweise sind die Eierstöcke sackartig und gepaart und haben sowohl in Teleostern als auch in Elasmobranchen eine symmetrische Struktur und sind funktionell. In einigen Elasmobranchen wird das rechte Ovar jedoch funktionsfähig, in einigen Fällen entwickelt sich nur das linke Ovar.
Histologie:
Der Eierstock enthält Stützgewebe, bekannt als Stroma, das Oogonia und Oozyten enthält. Das Oogonium (Keimzelle) entwickelt sich nach der Vermehrung zu einer primären Eizelle. Die Oogoniumzelle in den frühen Stadien der Entwicklung ist eine große Zelle mit einem großen Kern mit prominentem Nucleolus mit chromophobem Zytoplasma. Die Größe der Zelle wird nach der Multiplikation kleiner, ist aber nicht wichtig.
Der wichtige Unterschied besteht in den Chromosomen. Das Chromosom wird fadenartig im Zellkern (Leptoten-Stadium). Die Chromosomen durchlaufen das Zygotenen-Stadium, gefolgt von Pachyten und schließlich das Diplotene-Stadium. In diesem Stadium der Entwicklung ist die Eizelle von Follikelzellen umgeben. Die Follikelzellen sind in elasmobranch und amniota mehrschichtig, in teleosts jedoch einschichtig.
Bei Säugetieren bilden die follikulären Zellen ein geschichtetes Epithel, die die Eizelle umgebende Stratum granulosa, die sich schließlich zu Granulazellen vermehren. Die den Follikel unmittelbar umgebenden Stromazellen bilden eine Hülle aus Zellen, den Thekafollikel, der sich in Theca externa und Theca Interna unterscheidet.
In elasmobranch liegt die Theca interna unmittelbar unterhalb der Außenseite und besteht aus 4 bis 6 Reihen abgeflachter und langgestreckter Zellen, deren Wände sich zu Prozessen nach außen erstrecken (Dodd, 1983). Theca externa-Zellen haben eine Sekretion und enthalten Organellen wie ER und Mitochondrien.
Es enthält Fibroblasten, Kollagenfasern und Blutkapillaren sowie steroidproduzierende Zellen. Die reife Eizelle besteht aus zwei Schichten, einer äußeren Thekaschicht und einer inneren Granulosaschicht, die beide durch eine ausgeprägte Basalmembran voneinander getrennt sind (Abb. 20.4).

Der Eierstock enthält zahlreiche Oozyten in verschiedenen Entwicklungsstadien und Degenerationen. Die follikuläre Atresie ist ein degenerativer Prozess, bei dem Oozyten in verschiedenen Wachstums- und Differenzierungsstadien des Eierstocks des Fisches sowie der Eisprung verloren gehen. Follikel verloren ihr Wachstum und ihre Differenzierung. Sie synthetisieren keine Steroidhormone und üben keine endokrine Funktion aus.
Die dengerierenden Follikel sind zwei Arten, der Corpora lutea und der Corpora atretica. In Corpora lutea werden die eingeschlossenen Oozyten durch die eindringenden Granulosazellen und die Einwachsen von Thekazellen aus den Follikeln entfernt. Corpora lutea und Corpora atretica sind im Eierstock des Holocephalon vorhanden.
Bei Ei- und Vivipararten sind sie normalerweise im Spätsommer vorhanden. Sie stellen wahrscheinlich yolky Oozyten dar, die aus irgendeinem Grund nicht ovuliert werden konnten. In Squalus acanthias corpora atretica fehlt in den Eierstöcken.
Vitellogenese:
Das Ei am Anfang ist ohne Eigelb, aber die Vitellogenese findet später statt (Atresie). Bei oviparen und ovoviviparen Wirbeltieren ist seit langem bekannt, dass das Blut einer reifen Frau ein Kalzium bindendes Lipophosphoprotein, Vitellogenin, enthält, das in der Leber unter Stimulierung weiblicher Sexualsteroide synthetisiert wird.
Es gelangt aus dem Plasma in Oozyten, in denen es zu dem Eigelbprotein Lipovitellin und Phosvitin führt (Wallace, 1978). Die Vitellogenese im Hundefisch ist ähnlich wie bei anderen Wirbeltieren. Craik (1978) maß die Geschwindigkeit, mit der Vitellogenin synthetisiert und in Eigelbkörnchen umgewandelt wird.
Es findet die Vitellogenese statt, bei der eine Ansammlung von Dottersubstanz auftritt. Die auffällige Veränderung ist, dass das Cytoplasma, das basophil war, jetzt acidophil wird. Es gibt drei essentielle Dottersubstanzen - die Eigelbvesikel, die Eigelbkügelchen und Öltröpfchen.
Das Eigelb-Vesikel ist mit Glykoprotein versehen, das sich mit Eosin leicht rot färbt, aber PAS-positiv ist. Sie nehmen tiefrote Flecken. Die Dotterbläschen werden später zu kortikalen Alveolen und beteiligen sich an der Bildung des Perivitellinraums.
Die Eigelbkügelchen bestehen hauptsächlich aus Lipoprotein mit einigen Kohlenhydraten und anderen Substanzen. Sie sind für PAS schwach positiv. Die alten Tröpfchen enthalten im Allgemeinen Glyceride und eine geringe Menge Cholesterin. Mit der Vervollständigung der Vitellogenese-Bewegung der Keimvesikel kommt es zu einer Verschmelzung der Dottergranulate und zur Gruppierung von Öltröpfchen.
Nach der Bewegung der Keimbläschen zu einem Tierpol erfolgt die erste meiotische Teilung und der erste Polkörper wird freigesetzt.
Anschließend beginnt die zweite meiotische Teilung und ein Ei, in dem die an der Metaphase festgehaltene Teilung ovuliert wird. Chieffi; glaubt, dass echte Corpora lutea (dh im funktionalen Sinn von Säugetieren) sowohl in oviparen als auch in viviparen Elasmobranch gefunden wird. In ersterem (S. stellaris) entwickeln sie sich wahrscheinlich von ovulierten Follikeln (Corpora lutea) und in letzteren (T. marmorata) durch follikuläre Atresie (Corpora atretica).
Steroide sind im Eierstock von Elasmobranch vorhanden. Im Eierstock von Squalus waren 17 beta-Estradiol sowie Spuren von Progesteron und Östrogen vorhanden. In Squalus acanthias wird gezeigt, dass Progesteron aus (HC) Pregnenolon synthetisiert werden kann.
Die Enzyme, die für die Steroidogenese von Squalus acanthias erforderlich sind, wie Glucose-6-phosphatdehydrogenase (G-6PDH), 3 beta-Hydroxysteroiddehydrogenase (3 beta HSD), 3 alpha-Hydroxysteroiddehydrogenase (17 beta HSD) und 20 beta Hydroxysteroiddehydrogenase (20 Beta-HSD) sind im Eierstock vorhanden.
3-beta-HSD wird in Corpora lutea, aber nicht in Corpora atretica gefunden, und 3a-HSD war in den Granulosen der späten Corpora atretica schwach vorhanden.
Männliche Fortpflanzungsorgane:
Testes Morphologie:
Die Hoden bei Fischen sind im Allgemeinen gepaarte Strukturen, die auf beiden Seiten etwas seitlich und unterhalb der Nieren liegen. Der Hoden hat zwei Hauptfunktionen, die Produktion von Spermatozoen (Spermatogenese) und eine weitere Funktion ist die Produktion von Steroiden (Steroidogenese). Die Größe der Hoden in der Brutzeit wird enorm.
Die Hoden können länglich, sackartig oder lappig sein, können im Querschnitt abgerundet oder dreieckig sein. Bei reifen Gobies, Acanthogobius fluviatilis, ist der Hoden klein und fadenförmig. In Notopterus und amphipnous Hoden ist ungepaart. Bei Syngnathids ist der Hoden eine einfache Röhre. Die Hoden nehmen unterschiedliche Positionen in der Bauchhöhle ein.
Sie werden durch ein als Mesorchia bezeichnetes Mesenterium an der dorsalen Körperwand aufgehängt, das vaskularisiert ist und Nervenfasern enthält. Bei Dipnoifischen, z. B. bei Neoceratodus und Protopterus, ist der rechte Hoden anterior an der Spitze der Leber befestigt, während sich die linke Gonade bis zum Bereich des Ductus cusvieri erstreckt (Abb. 20.5).
Das Recht ist normalerweise nach links größer. Die Hoden des Protopterus erstrecken sich über die gesamte Körperlänge als dickes Band, das eng und breit an die Niere gebunden ist. Der Hoden von Lepidosiren ist eine langgestreckte Masse, die in Fettgewebe eingebettet ist.

Die Hoden von Teleostern sind lang und im Schnitt gerundet. In Latimaria ist der rechte Hoden zwei- bis dreimal größer als der linke, aber beide produzieren Spermien. Der rechte Hoden erstreckt sich durch die Mitte des ventrolateralen Körpers der Körperhöhle bis zur Schwimmblase.
In Chondrichthyes variiert die Position der Hoden von weit nach anterior bis etwa zur Mitte. Bei einigen Haien ist der Hoden mit einem epigonalen Organ verbunden, das von Natur aus lymphoid ist.
Aus jedem Hoden entsteht ein Samenleiter oder Samenleiter. Bei einigen Fischen vereinigen sich mesonephrische Gänge mit Hoden zu Vas deferens und Vasa Efferentia (epidydimous). Bei einigen Fischen wird der Spermiengang mit der Niere geteilt und oft als nephrischer Gang (Wolff'scher Gang) bezeichnet (Abb. 20.6a, b).

Bei Lungenfischen erstrecken sich viele efferente Gänge vom Zentralkanal in die Niere und sind mit den Kapseln der Nephrentubuli verbunden. Bei Teleostern ist der Spermiengang oder der Samenleiter ein modifizierter nephrischer Gang und kann von einem oder mehreren Harnleitern stammen, die die Ausscheidungsflüssigkeit zur Kloake transportieren (Abb. 20.6cd).
Der Samenkanal:
Der Spermiengang öffnet sich in eine andere Kammer, die Samenbläschen genannt wird. Das Samenbläschen ist verdickt und hat oft einen größeren Durchmesser als der Spermiengang. Die Vas deferens und Samenblasen sind bei den Haien deutlich sichtbar. Dort werden die Spermien für kurze Zeit gelagert.
Es wird einhellig angenommen, dass das Samenbläschen in Teleostern fehlt, weil es keine Spermien speichert und nicht Teil des Wolffschen Gangsystems ist. Daher ist es nicht homolog zu den Samenbläschen höherer Wirbeltiere. Bei Goldfischen bildet eine mittlere Wand des Spermiengangs jedoch einen unauffälligen Vorsprung, in dem die Spermien gelagert werden.
Diese Struktur wird als homolog zu den Samenbläschen angesehen, obwohl sie nicht glandulär ist. In Chondrostei, Holostei und Dipnoi ist ein Samenblasen wie der von Knorpelfischen zu beobachten. Der Spermiengang aus jedem Hoden verbindet sich häufig zu einem gemeinsamen Gang und öffnet sich durch die Genitalpore nach außen.
In Squalus vereinigen sich die nephrischen Gänge im Sinus urinus, der auch als Harnleiter dient und in die Kloake mündet. Von der Kloake aus werden die Spermatophoren (Spermienbündel) durch die Wirkung von Siphons gezwungen, wobei die unter der Haut liegenden Muskulaturbeutel sich von der Kloake aus durch die Rillen der Klasper in die Kloake der Frau erstrecken.
Bei einigen Haien tritt ein Rudiment des Mullerschen Ganges ein. Bei Muraenids mündet die Harnhöhle in die Harnblase, bei Lachs und Barsch in die Ausscheidungshöhle. In Lota mündet es in den Analrand. Es öffnet sich in die gemeinsame Anal- und Ausscheidungsöffnung in Synganitiden (dh in eine Kloake).
Auf der Basis von internen Modifikationen und Differenzierung von Keimgewebe oder deren Freisetzung in der zentralen Kavität von röhrenförmigen oder lobulären Formen. In Chondrichthyes bildet die Hodenwand sphärische Ampullen oder Follikel und nimmt keine röhrenartige Struktur an.
Während der Entwicklungsstadien beginnt die Ampullenbildung am ventralen freien Rand, wo neue Follikel keimen, und schreitet in Richtung der dorsalen Befestigung voran.
Zum Zeitpunkt der Zucht sind die reifen Ampullen durch enge Ductuli recti mit dem Zentralkanal verbunden, der entlang des angebrachten Randes der Gonade liegt. Die Ampullen sind mit Keimzellen versehen und durchlaufen in den Ampullen Spermatogenese und setzen Spermien durch enge Kanäle in die zentrale Kammer frei. Die Ampullen stammen aus der ampullogenen Zone.
In Teleost werden sowohl Ampullen- als auch Röhrenformen beobachtet. Die Struktur von Ampullen ist der von Elasmobranchen ähnlich. Die röhrenartige Struktur reicht von der äußeren Tunica propria bis zum zentralen Hohlraum. Die Keimzellen oder Spermatogonien befinden sich in diesen röhrenartigen Anordnungen.
Die Keimzellen werden direkt in die zentrale Höhle abgegeben, da kein lobuläres Lumen vorhanden ist. In dieser Form gibt es üblicherweise einen zentralen Hohlraum, in den radial angeordnete kurze Tubuli münden. Die Hodenstrukturen finden sich in Guppy, Poe-cilia reticulata (Abb. 20.7a, b).

In Dipnoans befinden sich in Protopterus Halstubuli und in Lepidosiren Ampullen. Die seminiferen Tubuli sind viel größer als Ampullen von Lipidosiren. Die kanalikulären oder Rate-Erweiterungen der rechten und linken Hoden vereinigen sich zu einem Medianrohr.
Histologie:
Jeder Hoden ist in einer Bindegewebshülle eingeschlossen. Die Tunica Propria des Bindegewebes ragte in die Lumen bildenden Schläuche hinein. Das blinde Ende ist der Ort der primären Spermatozyten. Die röhrenförmige Struktur besteht aus zwei Teilen, dem Interstitial und dem lobulären. Der interstitielle Teil besteht aus interstitiellen Zellen, Fibroblasten sowie Blut- und Lymphgefäßen.
Der lobuläre Teil unterscheidet sich in Keimzellen und Körperzellen. Die somatischen Zellen sind positiv für Lipid und Cholesterin und sind homolog zu den Säuger-Leydig-Zellen. Einige Forscher gaben an, dass Cholesterin positive Lipide.
Die Art der Zelle ist nicht nur das Kriterium für ihre Homologie mit Leydig-Zellen, dass sie steroidproduzierende Zellen sind. Die Leydig-Zellen sezernieren Steroidhormone. Ein anderer Zelltyp im Interstitium sind Sertoli-Zellen. Bei den Sertoli-Zellen handelt es sich im Wesentlichen um säulenartige Zellen mit komplexen apikalen und lateralen Prozessen (Abb. 20.8), die von den Spermatogonzellen umgeben sind und den Raum dazwischen ausfüllen. Sie versorgen die Keimzellen mit Nährstoffen.
Bei Säugetieren kann seine Funktion das Sekretieren von Androgenbindungsprotein umfassen, das dazu dient, Testosteron innerhalb des semiferhaltigen Epithels und im proximalen Teil des Genitalgangsystems zu konzentrieren. Seine Sekretion hilft bei der Regulierung von Mitose, Meiose und steroidogenen Funktionen der Leydig-Zellen und der Freisetzung von Gonadotropinen aus der Hypophyse.

Grier und Linton 1977 fanden Sertoli-Homologezellen, während sie Sudan-Schwarzfärbung erhalten. In Bezug auf die Terminologie, ob diese Zellen Sertoli-Zellen, Zystenzellen oder Sertoli-homologe Zellen genannt werden sollten, besteht die Einigkeit, dass sie in Teleosts als Sertoli-Zellen bezeichnet werden können.
Spermatogenese:
Spermatogenese ist der Entwicklungsprozess, durch den die spermatogonischen Zellen von undifferenzierten diploiden (2n) Spermatogonien in hoch spezialisierte haploide (n) Spermatozoen umgewandelt werden.
Es gibt im Allgemeinen drei Hauptphasen:
(i) Die spermatogoniale Phase oder Spermatozytogenese.
(ii) Die meiotische Phase oder Meiose.
(iii) Spermatidphase oder Spermiogenese.
Das Spermatogonium im Frühstadium bei Fischen ist eine große ovale Zelle mit einem großen runden Nucleolus. Es wird in primäre Spermatozyten, dann in sekundäre Spermatozyten durch Reifung (oder Meiose) unterteilt und entwickelt sich schließlich zu Spermatiden. Die Spermatide entwickelt sich dann zu einem Spermatozoon.