Organische Evolution: 9 Hauptbeweise der organischen Evolution
Zur Unterstützung der organischen Evolution sind 9 wichtige Beweise wie folgt:
1. Paläontologische Beweise (Beweise aus Fossilienfunden):
Aus den Fossilienfunden wurde geschlussfolgert, dass die Entwicklung von einfach zu komplex schrittweise erfolgte.

Bildhöflichkeit: upload.wikimedia.org/wikipedia/commons/rex_p1050042.jpg
Zur Unterstützung werden im Folgenden einige Beweise gegeben.
(i) Anzahl und Art der Fossilien in frühen Gesteinen:
Die Gesteine der frühen Epoche (z. B. Proterozoikum) enthalten weniger Fossilien als die Gesteine der späteren Epoche, und in diesen Gesteinen befinden sich nur Fossilien von einfachen wirbellosen Meerestieren. Es ist darauf zurückzuführen, dass das Leben zunächst als einfache Form im Meer entstand. Fossilien waren also anfangs nicht in ausreichendem Maße vorhanden, wie sie später waren.

(ii) Verteilung von Fossilien in den aufeinanderfolgenden Schichten:
Die Verteilung der Fossilien deutet darauf hin, dass frühe Fossilien in den unteren Gesteinen einfach sind, die jüngsten Fossilien in den oberen Gesteinsschichten sind jedoch komplexer. Es zeigt, dass die fossilen Formen von Anfang an bis zu den jüngsten Gesteinen immer komplexer werden. Die Gesteine des Proterozoikums enthalten nur wenige Fossilien.
Das Paläozoikum enthält reichlich Fossilien von Wirbellosen, Fischen und Amphibien. Die Gesteine des Mesozoikums haben die Fossilien großer Reptilien (Dinosaurier) und primitiver Vögel und Säugetiere. Brontosaurus war ein Dinosaurier von Pflanzenfressern, 25 Meter lang, 4, 5 Meter hoch und 45 Tonnen schwer. Tyrannosaurus war der größte fleischfressende Dinosaurier, 5, 4 Meter hoch und 13 Meter lang. In der Koenozoik sind die Fossilien verschiedener Säugetiere reichlich vorhanden.
(iii) Ungleichheit zwischen den vergangenen und gegenwärtigen Lebensformen:
Auf der Grundlage fossiler Studien hat sich gezeigt, dass die frühen Organismen sich sehr von ihren modernen Formen unterschieden, dh der frühe Mensch lebte in den Höhlen ohne soziales Leben und verbrachte sein Leben wie Tiere, aber der Mensch machte Fortschritte und das Modem Der Mensch ist zivilisiert und führt ein starkes soziales Leben. So haben sich die Organismen seit ihrem Erscheinen verändert, was die Entwicklung der Evolution unterstützt.
(iv) Fehlende Links (Übergangsformulare):
Die fossilen Organismen, die Charaktere zweier verschiedener Gruppen zeigen, werden als fehlende Verbindungen bezeichnet.
Beispiele:
(a) Archaeopteryx (Archae - primitiv, alt, Pteryx = Flügel):
Es wurde in den Felsen der Jurazeit gefunden. Archaeopteryx lithographica wurde 1861 von Andreas Wagner aus dem lithographischen Steinbruch im bayerischen Solenhofen entdeckt. Dieses Fossil befindet sich im British Museum, London. Es zeigt die Charaktere sowohl der Reptilien als auch der Vögel.
Reptilianische Charaktere des Archaeopteryx:
(a) Die Körperachse ist mehr oder weniger eidechsenartig.
(b) ein langer Schwanz ist vorhanden,
(c) die Knochen sind nicht pneumatisch,
(d) Die Backen sind mit ähnlichen Zähnen versehen.
(e) Die Hand trägt einen typischen Reptilienplan und jeder Finger endet in einer Klaue.
(f) Vorhandensein eines schwachen Brustbeins
(g) Vorhandensein von freien Schwanzwirbeln, wie sie in Eidechsen zu finden sind.

Vogelfiguren des Archaeopteryx:
a) Vorhandensein von Federn am Körper,
(b) Die beiden Kiefer sind zu einem Schnabel umgebaut.
(c) Die Vorderglieder sind zu Flügeln modifiziert.
(d) Die Hinterbeine sind auf dem typischen Vogelplan aufgebaut.
(e) Eine intime Fusion der Schädelknochen, wie sie bei den Vögeln zu sehen ist.
Aus den obigen Tatsachen geht hervor, dass die Vögel aus Vorfahren der Reptilien entwickelt wurden. So ist Huxley berechtigt, "Vögel sind die verherrlichten Reptilien" zu nennen.
(b) Ichthyostega:
Es ist eine primitive fossile Amphibie und eine fehlende Verbindung zwischen Fischen und Amphibien.

(c) Seymouria:
Es war eine "fehlende Verbindung" zwischen Amphibien und Reptilien.
(d) Lycaenops:
Es war ein säugetierartiges Reptil. Es wird als "fehlende Verbindung" zwischen Reptilien und Säugetieren betrachtet.

(e) Cynognathus (Hundekiefer):
Es war ein säugetierähnliches Reptil und hatte sowohl Reptilien als auch Säugetiere. Es war einer der alten Vorfahren der Reptilien von Säugetieren.
(f) Basilosaurus:
Dieser fossile Wal hatte Hinterbeine. Es verbindet die Wassersäugetiere mit ihren irdischen Vorfahren.
(g) Pteridosperms (Cycadofilicales, Samenfarne):
Dies sind fossile Pflanzen, die zwischen Farnen und Samenpflanzen liegen.
Es gibt mehr Tierfossilien als Pflanzen. Es ist auf langsame, zerfallende härtere Strukturen im Endoskelett und Exoskelett zurückzuführen.
(v) Vorfahren einiger Tiere:
Paläontologen haben aus den Studien ihrer Fossilien vollständige Entwicklungsgeschichten einiger Tiere wie Pferde, Kamel, Elefanten und Menschen abgeleitet.
Entwicklung des Pferdes:
Die Entwicklung des Pferdes wurde 1879 von Othniel C. Marsh beschrieben. Herkunftsort - Der Ursprung des Pferdes fand im Eozän statt. Das erste Fossil des Pferdes wurde in Nordamerika gefunden. Es wurde Eohippus genannt, aber später in Hyracotherium umbenannt.

Evolutionärer Trend:
Die ständige Veränderung eines Charakters innerhalb einer sich entwickelnden Linie wird als evolutionärer Trend bezeichnet. Eine Linie ist eine evolutionäre Sequenz, die in linearer Reihenfolge von einer Vorfahrengruppe zu einer Nachkommengruppe angeordnet ist. Ein Trend kann progressiv sein (allgemeine Zunahme der Organgröße) oder rückläufig (allgemeine Degeneration und Verlust von Organen).
Die folgende Liste zeigt den wichtigsten evolutionären Trend von Pferden.
(a) Vergrößerung,
b) Dehnung von Hals und Kopf
c) Verlängerung der Vorder- und Hintergliedmaßen
(d) Verringerung der seitlichen Ziffern,
(e) Zunahme der Länge und Dicke der dritten Ziffer,
(f) Begradigung und Versteifung des Rückens
(g) Vergrößerung und Komplexität des Gehirns
h) besser entwickelte Sinnesorgane,
(i) Zunahme der Zahnlänge
(j) Zunahme der Breite der Schneidezähne
k) Ersatz von Prämolaren durch Molaren.
(l) Erhöhung der Kronenhöhe der Molaren,
(m) erhöhte seitliche Unterstützung der Zähne durch Zement,
(n) Vergrößerte Oberfläche der Höcker durch die Entwicklung von Schmelzrippen (Änderung der Prämolaren und Molaren von der Art der Färbung zu der Art der Weide).
Die Evolution des modernen Pferdes wird kurz wie folgt beschrieben:
Eohippus (= Hyracotherium):
Die Entwicklung des modernen Pferdes begann vor etwa 60 Millionen Jahren in der Eocine-Epoche. Wie bereits erwähnt, wurde in Nordamerika das erste Fossil namens Eohippus, 'Dawn Horse', gefunden. Dieses Pferd war etwa so groß wie ein Fuchs- oder Terrierhund (eine Art kleinhaariger Hund zum Aushängen von Füchsen) und nur 30 cm hoch an den Schultern. Es hatte kurzen Kopf und Hals.
Die Vorderfüße hatten vier vollständige Finger (2, 3, 4 und 5) und eine Schiene des ersten Fingers und die Hinterfüße mit drei funktionellen Zehen (2, 3 und 4) und zwei Schienen der ersten und fünften Zehen. Schienen sind reduziert und nicht funktionelle Seitenfinger und Zehen des Pferdes. Die Zähne befanden sich in vollständigem Zement. Molare Zähne hatten keine Zähne. Niedrig gekrönte Backenzähne wurden für das Durchsuchen der weichen üppigen Vegetation angepasst.
Orohippus:
Seine Fossilien stammen aus dem mittleren Eozän. Es war etwas höher als Eohippus. Seine Vorderfüße behielten vier Finger, und die Hinterfüße hatten drei Zehen, aber kleine Reste der beiden Zehen in den Hinterfüßen gingen verloren. Die mittlere Ziffer in beiden Gliedmaßen war dominant. Die dritten und vierten Prämolaren zeigten einen Trend zum Molaren, aber das Tier war immer noch ein Browser.
Mesohippus:
Mesohippus, das Zwischenpferd, entwickelte sich vor etwa 40 Millionen Jahren während der Oligozän-Epoche aus Hyracotherium. Es war etwa so groß wie moderne Schafe an den Schultern. Vier Fuß hatten drei Finger und eine Schiene des fünften Fingers, und die Hinterfüße besaßen drei Zehen, aber der mittlere war länger als andere und stützte das meiste Körpergewicht. Backenzähne hatten einige Zacken.
Miohippus:
Im späten Oligozän wurde Mesohippus durch Miohippus ersetzt. Es war dem Mesohippus sehr ähnlich, aber etwas größer. Obwohl beide Fuß- und Hinterfüße drei Zehen waren, waren die Zehen breit und breiteten sich aus. Dies deutete darauf hin, dass Miohippus auch Waldbewohner war. Die Zähne waren noch tief gekrönt und es war immer noch ein Browser.
Parahippus:
Es stammt von Miohippus in der frühen Miozän-Epoche ab. Seine Gliedmaßen hatten immer noch drei Ziffern, zeigten jedoch eine deutliche Dehnung. Parahippus hatte Hypsodontenzähne (Zähne mit offenen Wurzeln und hartnäckigen Pulpen, so dass sie, so schnell sie abgenutzt sind, weiter wachsen).
Merychippus:
Merychippus, das wiederkäuende Pferd, entstand vor 25 Millionen Jahren aus Mesohippus im Miozän. Es hatte die Größe eines kleinen Ponys und war an den Schultern etwa 100 cm hoch. Es war mit längerem Hals. Seine Vorder- und Hinterfüße hatten drei Finger und drei Zehen, wobei der Mittelfinger und die Zehe länger als andere waren und das gesamte Körpergewicht stützten. Es gab keine Schiene. Zähne waren länger mit Zement. Molare Zähne hatten gut entwickelte Zähne.
Pliohippus:
Pliohippus, das Pliozänpferd, entstand vor etwa 10 Millionen Jahren aus Merychippus in der Pliozän-Epoche. Es war die Größe eines modernen Ponys, etwa 120 cm hoch an den Schultern. Jeder Vorder- und Hinterfuß hatte einen vollständigen Finger und einen kompletten Zeh und zwei unter der Haut verborgene Schienen. Pliohippus wird daher als das erste ein Pferd bezeichnet. Die Backenzähne waren lang mit gut entwickeltem Zement und Kerbverzahnungen. Zähne wurden für das Essen von Gras angepasst.
Equus:
Dies ist das moderne Pferd, das vor neun bis zehn Jahren in Nordamerika aus Pliohippus in der Pleistozän-Epoche hervorging und später mit Ausnahme Australiens auf der ganzen Welt verbreitet wurde. Sie ist an den Schultern etwa 150 cm hoch. Es hat einen langen Kopf und einen langen Hals. Jeder Vorder- und Hinterfuß des modernen Pferdes hat einen Finger, einen Zeh und zwei Schienen. Die Kronen der Backenzähne sind mit emaillierten Stegen verlängert und eignen sich hervorragend zum Schleifen.
2. Beweise aus vergleichender Anatomie und Morphologie:
Es gibt Ähnlichkeiten und Unterschiede zwischen Organismen von heute und solchen, die vor Jahren existierten. Diese Beweise sind wie folgt.
(i) Organsysteme:
Die verschiedenen Systeme des tierischen Körpers sind in vielen Organismengruppen ähnlich, z. B. Nervensystem, Blutgefäßsystem, Atmungssystem, Ausscheidungssystem usw. Das Atmungssystem von terrestrischen Wirbeltieren hat zwei Lungen, eine Trachea, einen Kehlkopf, Nasenkammern und Nasenlöcher . Ebenso enthält das Blutgefäßsystem aller Wirbeltiere ein Herz, Arterien, Venen und Lymphgefäße.
Transportsysteme von Pflanzen haben ähnliche Arten von leitenden Kanälen aus Xylem und Phloem. Das Vorhandensein ähnlicher Organsysteme weist auf eine gemeinsame Abstammung hin. Trotz der großen Ähnlichkeit weisen die Organsysteme verschiedener Gruppen je nach Lebensraum und Ausmaß der Evolution unterschiedliche Spezialisierungen auf.

(ii) homologe Organe:
Richard Owen (1804-1892) führte den Begriff homolog ein. Die Organe, die dieselbe Grundstruktur haben, aber unterschiedliche Funktionen haben, werden als homologe Organe bezeichnet. Diese Organe folgen während ihrer Entwicklung demselben Organisationsplan. Im erwachsenen Zustand werden diese Organe jedoch so modifiziert, dass sie unterschiedliche Funktionen als Anpassung an unterschiedliche Umgebungen erfüllen. Die homologen Strukturen sind das Ergebnis einer unterschiedlichen Entwicklung. Homologie weist auf gemeinsame Abstammung hin.
Beispiele:
(a) Die Vorderglieder des Menschen, des Geparden, des Wals und der Fledermaus haben den gleichen Grundstrukturplan. In jedem Fall besteht das Vorderbein aus Humerus, Radioulna, Karpalen, Metacarpalen und Digits. Die Skelettteile der Vorderbeine aller dieser Wirbeltiere sind in Aufbau und Anordnung ähnlich. Die Vorderglieder dieser Tiere haben jedoch unterschiedliche Formen und Funktionen. Beim Menschen werden sie zum Greifen verwendet, beim Geparden zum Laufen, beim Wale zum Schwimmen und beim Fledermaus (1. 7.19).

(b) Eine strukturelle Homologie wird auch im Skelett, Herz, Blutgefäßen, Gehirn, Nerven, Muskeln und Ausscheidungssystem verschiedener Wirbeltiere beobachtet.

(c) Ein anderes Beispiel für homologe Organe sind verschiedene Mundteile einiger Insekten. Die Maulteile von Kakerlake, Honigbiene, Mücke und Schmetterling haben den gleichen Grundplan. Bei jedem dieser Insekten bestehen die Maulteile aus Labrum, einem Paar Mandibeln und zwei Paaren von Maxillen, die jedoch unterschiedliche Funktionen erfüllen, wobei sie ihre unterschiedlichen Ernährungsgewohnheiten im Blick behalten. Die Maulteile der Kakerlake sind zum Beißen und Kauen geeignet. In Honigbiene zum Kauen und Läppen, in Mücke zum Stechen und Saugen, in Stubenfliege zum Schwämmen und in Schmetterling zum Abtropfen.
(d) In Pflanzen können die homologen Organe ein Dorn von Bougainvillea oder eine Ranke von Curcurbita sein, die beide in der Achselhöhle auftreten. Blätter höherer Pflanzen entstehen aus Knoten, besitzen Achselknospen und erzeugen eine Lücke in der Gefäßversorgung des Stiels. In der Form können sie sitzend (z. B. Zinnia) oder gestielt (z. B. Pipal), einfach (z. B. Mango) oder Verbindung (z. B. Cassia) sein und zu Schuppen (z. B. Spargel), die zu Stacheln (z. B. Berberitze) modifiziert sind, reduziert sein. zum Schutz und Ranken (zB Lathyrus aphaca) zum Klettern. Die Modifikationen zeigten die Entwicklung des Organs an die verschiedenen Funktionen an.

(e) Homologie ist auch unter den Molekülen zu sehen. Dies wird als molekulare Homologie bezeichnet. Zum Beispiel sind die im Blut von Menschen und Affen gefundenen Proteine ähnlich. Die Phylogenie eines Organismus kann unter Verwendung der Basensequenz in Nukleinsäuren und der Aminosäuresequenz von Proteinen in verwandten Organismen verfolgt werden.

(iii) analoge Organe:
Organe, die ähnliche Funktionen haben, sich aber in ihren strukturellen Details und ihrem Ursprung unterscheiden, werden als analoge Organe bezeichnet. Die analogen Strukturen sind das Ergebnis konvergenter Evolution.
Beispiele:
(a) Die Flügel eines Insekts entsprechen den Flügeln eines Vogels. Dies ist darauf zurückzuführen, dass sich die Grundstruktur der Flügel der Insekten von den Flügeln des Vogels unterscheidet. Ihre Funktion ist jedoch ähnlich (Abb. 7.23).
(b) Brustflossen von Haien und Delfinenflossen sind analoge Organe. Brustflossen von Haien sind nicht pentadaktil. Die Flossen der Delphine sind Pentadactyle. Die Grundstruktur der Brustflossen von Haien und der Flossen von Delphinen ist jedoch unterschiedlich, aber beide sind beim Schwimmen nützlich (Abb. 7.24).
(c) Stiche von Honigbiene und Skorpion sind analoge Strukturen. Der Stachel der Honigbiene ist eine Modifikation des Ovipositors (Struktur, die bei der Eiablage hilft), während der Skorpion das letzte Abdomensegment modifiziert. Stiche beider Arthropoden haben eine ähnliche Funktion.

(d) Blätter sind auf die Photosynthese spezialisierte Pflanzenorgane. Es gibt jedoch Pflanzen, bei denen die Blätter als Reaktion auf eine bestimmte Umgebung oder Anforderung modifiziert oder reduziert werden. Die Funktion der Blätter übernehmen dann andere Organe wie Nebenblätter (z. B. Lathyrus aphaca), Blattstiel (z. B. Acacia auriculiformis) und Stieläste (z. B. Ruscus, Spargel). Sie sind unter ihnen genauso wie bei den Blättern.
Ebenso sind Pflanzenranken zum Klettern bestimmt. Sie können von Stängeln (zB Passiflora), Blättern (zB Lathyrus aphaca, Pisum sativum) abgeleitet sein. Das Vorhandensein analoger Organe weist auf eine ähnliche Anpassung durch nicht verwandte Gruppen durch Modifikation oder Entwicklung verschiedener Teile hin. Es wird konvergente Evolution genannt.

(iv) Überflussorgane:
Die Organe, die in reduzierter Form vorliegen und keine Funktion im Körper haben, sondern den vollentwickelten Funktionsorganen verwandter Tiere entsprechen, werden als Überrestorgane bezeichnet. Es wird vermutet, dass es sich dabei um Überreste von Organen handelt, die bei ihren Vorfahren vollständig und funktionsfähig waren.

Beispiele:
(a) Überflussorgane im menschlichen Körper:
Es wurde beschrieben, dass der menschliche Körper etwa 90 Restenorgane besitzt. Einige davon sind Nictitating (plica semilunaris) -Membran, Ohrmuskulatur (Muskeln der Ohrmuschel), Segmentmuskeln des Unterleibs, Panniculus camosis (Unterhautmuskulatur), Wurmfortsatz, Caudalwirbel (auch Steißbein oder Schwanzbein genannt), dritte Molaren (Weisheit) Zähne), Haare am Körper und Brustwarzen bei Männern (Abb. 7.26).
(b) Vestigialorgane bei Tieren:
Wichtige Beispiele sind Überreste von Hinterbeinen und Beckengürteln von Pythons (Abb. 7.27) und Grönlandwale (die zeigen, dass sich Schlangen und Wale ursprünglich aus vierbeinigen Vorfahren entwickelten), Flügel von flugunfähigen Vögeln wie Ostrich; Emu, Cassowary, Kiwi, Rhea und Dodo (ausgestorben), Schienenknochen in Pferdefüßen und Stirnfleck im Froschkopf (Überrest des 3. Auges).

c) Überlebensorgane in Pflanzen:
Eine oder mehrere Staminoden (Vestigial Staubblätter) kommen in den Blüten von mehreren Pflanzen vor, die zu Labiatae, Scrophulariaceae, Casesalpinioideae, Cucur-bitaceae usw. gehören. Nicht-fiktive Stempeln, genannt Pistilloides, treten in den männlichen Blüten der Cucurbitaceae auf.
In den Strahlenblüten von Sunflower fehlen die Staubblätter, während der Stempel rudimentär ist und ein kleines funktionsloses Stigma und einen ovulationslosen Eierstock aufweist. Blätter werden in Cuscuta, Orobanche, Spargel, Ruscus und einer Reihe anderer Pflanzen auf Schuppen reduziert.
(v) Verbindungslinks:
Die Organismen, die die Charaktere zweier verschiedener Gruppen besitzen, werden als Verbindungsglieder bezeichnet. Im Folgenden sind einige wichtige Beispiele für Verknüpfungen aufgeführt.
Beispiele: (a) Euglena ist ein Chlorophyll, das grünes Protozoon enthält und das Bindeglied zwischen Tieren und Pflanzen bildet.
(b) Proterospongie ist ein koloniales Protozoon. Es besteht aus Flagellaten und Collared-Individuen, die an Choocyten (Halszellen) von Schwämmen erinnern. Somit ist es eine Verbindung zwischen Protozoen und Porifera.

(c) Neopilina (Abb. 7.29). Es ist eine Verbindung zwischen Annelida und Mollusca. Es ähnelt Mollusken, da es eine Schale, einen Mantel und einen großen muskulösen Fuß besitzt. Zu den anneliden Charakteren gehören segmental angeordnete Kiemen, Nephridien und Muskeln sowie ein trochophorähnliches Larvenstadium.

(d) Peripatus (Abb. 7.30), eine Arthropode, ist ein Bindeglied zwischen Annelida und Arthropoda. Zu den Arthropoden-Charakteren zählen Hämocoel, Tracheae als Atmungsorgane und röhrenförmiges Herz mit Ostien. Die ausgestellten Annelid-Charaktere sind der wurmartige Körper, die Struktur der Augen, die unzusammenhängenden Beine, das Vorhandensein von segmentalen Nephridien, die weiche Kutikula und durchgehende Muskelschichten in der Körperwand.
(e) Balanoglossus. Es ist ein Hemichordat (Nichtchordat) und ist ein Bindeglied zwischen Nichtchordaten und Chordaten.
(f) Die Lungenfische, z. B. Protopterus (afrikanischer Lungenfisch), Lepidosiren (südamerikanischer Lungenfisch) und Neoceratodus (australischer Lungenfisch), können als Verbindungsglied zwischen den Fischen und Amphibien angesehen werden. Die Lungenfische haben alle Charaktere eines typischen Fisches, aber sie können durch die Lunge atmen und besitzen ein dreikammeriges Herz.

g) Latimeria (Coelacanth-Fische) gilt als Bindeglied zwischen Fisch und Amphibien.
(h) Chimaera. Es ist ein Bindeglied zwischen Knorpelfischen und Knochenfischen.
(i) Eier legende Säugetiere (z. B. Ornithorhychus, Entenschnabeltier und Tachyglossus oder Echidna oder Stacheliger Ameisenfresser) tragen Haare und Brustdrüsen, besitzen aber auch einige reptilische Merkmale wie das Legen von Eiern, das Vorhandensein von Cloaca und einige Skelettähnlichkeiten. Sie sind somit ein Bindeglied zwischen Reptilien und Säugetieren.

(vi) Atavismus:
Es ist das Wiederauftauchen bestimmter Ahnenfiguren, die entweder verschwunden waren oder reduziert wurden. Es gibt einige Beispiele für Atavismus beim Menschen, nämlich die Fähigkeit, bei einigen Personen die Ohrmuschel zu bewegen, stark entwickelte Eckzähne, außergewöhnlich lange, dichte Haare, bei manchen Babys kurzer Schwanz und bei manchen Individuen zusätzliche Säugetiere.
Atavismus wird auch in Pflanzen beobachtet. Bei Zitrusblättern ist die Lamina durch ein Gelenk oder eine Verengung vom Flügelstiel getrennt. Manchmal ist der geflügelte Teil des Blattstiels vergrößert, so dass zwei seitliche Blättchen entstehen, die das Blatt trifoliolieren.
Es zeigt, dass Citrusblatt einmal eine Trifoliolatverbindung war, aber während der Evolution sind zwei Blättchen degeneriert. In vielen Pflanzen (z. B. Rosa, Hibiscus, Oxalis, Poppy) werden einige der Staubgefäße und sogar der Fruchtblätter in blütenblattartige Strukturen umgewandelt, was darauf hindeutet, dass sich Staubgefäße und Fruchtblätter aus blattähnlichen Strukturen entwickelt haben.

3. Embryologische Beweise (Beweise aus der Embryologie):
Diese Nachweise basieren auf der vergleichenden Studie der Embryonen verschiedener Tiere.
(i) Ähnlichkeit in der frühen Entwicklung:
Bei allen mehrzelligen Tieren wird das befruchtete Ei (Zygote) segmentiert (Spaltung), um eine feste Struktur, die Morula, zu erzeugen. Die Morula entwickelt sich zu einer einschichtigen hohlen Blastula. Die letztere verwandelt sich in eine zwei- oder dreischichtige Gastrula. Die Tiere mit zweischichtiger Gastrula sollen diploblastisch sein, z. B. Coelenterate.
Die Tiere, bei denen dreischichtige Gastrula gefunden wird, sind als Triploblasten bekannt, wie Frosch, Eidechse usw. Diploblastische Gastrula besteht aus Ektoderm und Doderoderm: Diese zwei oder drei Schichten der Gastrula werden als primäre Keimschichten bezeichnet, die Anlass geben das ganze Tier. Eine solche ähnliche frühe Entwicklung stellt eine enge Beziehung zwischen allen mehrzelligen Tieren her.
(ii) Ähnlichkeit zwischen Wirbeltierembryonen:
Wenn eine vergleichende Studie von Embryonen im gleichen Alter von Wirbeltieren wie einem Fisch, einem Salamander, einer Schildkröte, einem Küken und einem Mann durchgeführt wird, wird beobachtet, dass sie sich sehr ähneln (Abb. 7.35). Sie haben mehr oder weniger die gleiche Form und Struktur wie Kiemenspalten, Schwanz usw. Obwohl die Embryonen aller Wirbeltiere einander ähneln, ähneln sich die Embryonen eng verwandter Gruppen stärker als die Embryonen der entfernten Gruppen. Dies ist ein weiterer Beweis für eine enge Beziehung zwischen diesen unterschiedlichen Wirbeltieren.
(iii) Ähnlichkeiten zwischen wirbellosen Larven:
Anneliden und Weichtiere besitzen eine ähnliche Art von Larve, die als Trochophor bezeichnet wird. Echinodermen und Hemichordate haben auch ähnliche Larven. Larvalähnlichkeit weist auf eine gemeinsame Abstammung hin.

(iv) progressive Metamorphose:
Die Ammocoeten-Larve von Lamprey ähnelt in den meisten Einzelheiten der erwachsenen Form von Amphioxus oder Branchiostoma, die nur möglich sind, wenn wir annehmen, dass Lamprey sich wie Tiere von Branchiostoma entwickelt hat.
(v) retrogressive Metamorphose:
Tiere wie Sacculina und Manteltiere (z. B. Herdmania) sind entartet und weisen keine Ähnlichkeit mit anderen Tiergruppen auf. Das Studium ihrer Embryologie hat jedoch geholfen, aufgrund der in ihren Embryonen vorhandenen Charaktere ihre wahre systematische Position zu finden. Sacculina ist ein Parasit auf Krabben.
Gliedmaßen, Mund, Verdauungskanal und spezielle Sinnesorgane fehlen. Der Parasit hat einen gestielten ovalen Sack, der zur Aufnahme von Nahrung Auswuchs in den Wirt schickt. Die taxonomische Position von Sacculina wurde durch das Studium seiner Larve, die der Nauplius-Larve von Krebstieren ähnelt, festgestellt.
In ähnlicher Weise hat die Tunika Herdmania einen einfachen, geldbeutelähnlichen Körper, der keine Spur für die Akkordation zeigt. Seine Larve besitzt jedoch alle wichtigen Eigenschaften der Chordate, was beweist, dass Herdmania eine Chordate ist.
(vi) Temporäre embryonale Strukturen:
Embryonen besitzen oft Strukturen, die bei Erwachsenen nicht vorkommen. Zum Beispiel hat Vogelembryo Zahnknospen und Kiemenspalten, die im erwachsenen Tier nicht gefunden werden. Das Vorhandensein von Zahnknospen hat keine Bedeutung für den Embryo, da die Nahrung aus Eigelb durch spezielle Blutgefäße gewonnen wird. Der Erwachsene, der sich von harten Körnern und Samen ernährt, braucht die Zähne, ist aber frei von ihnen.
Das Vorhandensein von Zahnknospen in den Embryonen kann nur unter der Annahme erklärt werden, dass
(i) Vögel haben sich aus gezahnten Vorfahren entwickelt;
(ii) Vögel haben während der Evolution Zähne verloren;
(iii) Der Vogelembryo besitzt aufgrund der Persistenz einiger Gene, die seine Wirkung während der Entwicklungsstadien ausdrücken, einige Vorfahren.
Wal ist ein Wassersäugetier. Es besitzt keine Körperbehaarung. Sein Fötus oder Embryo besitzt Haare, die vor der Geburt abgeworfen werden. Haare sind für den Embryo nutzlos, weil er im Körper der Mutter gut geschützt ist. Der frühe Froschkatt besitzt Kiemen und Schwanz, während der Metamorphose verschwinden diese Strukturen.
(vii) Entwicklung von Wirbeltierorganen:
Die Entwicklung vieler Organe von Wirbeltieren (z. B. Herz, Gehirn, Niere) zeigt den möglichen Weg der Evolution sowie die gemeinsame Abstammung von Wirbeltieren an. Zum Beispiel wird das Herz eines Säugetiers oder eines Vogels während seiner Entwicklung anfangs zwei Kammern (wie bei Fischen), dann drei Kammern (wie bei Amphibien und einigen Reptilien) und schließlich vier Kammern. Es zeigt deutlich, dass Vögel und Säugetiere von Fischen über Amphibien und Reptilien stammen.
Bei allen Wirbeltieren entsteht das Gehirn als vordere Vergrößerung des Neuralrohrs. Bald entwickelt er zwei Furchen und wird in drei Teile geteilt - Vorderhirn, mittleres und Hinterhirn. Jeder dieser Teile entwickelt sich weiter, um den Erwachsenenzustand zu erreichen.
Wirbeltiere haben drei Arten von Nieren: pronephrische, mesonephrische und metanephrische. Pronephrische Niere kommt bei Hagebutten vor. Mesonephrische Nieren kommen in anderen Fischen und Amphibien vor, während metanephrische Nieren in Reptilien, Vögeln und Säugetieren vorkommen. Im Säugetier- oder Vogelembryo ist die Niere anfangs pronephrisch, dann mesonephrisch und letztendlich metanephrisch.
(viii) Nachweise von Pflanzenembryonen:
(a) In Pinus treten die Laubblätter nicht direkt auf den Hauptstielen auf, sondern werden in Gruppen auf den Zwergentrieben getragen. Im Sämlingszustand treten die Laubblätter jedoch direkt auf dem Hauptstamm auf, was auf die Entwicklung von Pinus von Vorfahren hindeutet, die Laubblätter direkt auf den Hauptstielen besaßen.
(b) Australische Akazienarten besitzen Phylloden (Abb. 7.36) oder Blatt-Blattstiele anstelle von normalen zweipinnigen Blättern wie bei anderen Akazienarten. Australische Arten zeigen alle Übergangsstufen zwischen zweizähnigen Blättern und Phylloden während des Keimlingsstadiums.
(c) Viele Bryophyten durchlaufen ein Stadium filamentöser Protoneme, bevor sie die Erwachsenenform erreichen. Das filamentöse Protonem lässt auf Algenstämme von Bryophyten schließen.
(d) Bryophyten und Pteridophyten hatten männliche Gameten oder Spermien. Sie benötigen eine externe Wasserquelle, um zu den weiblichen Geschlechtsorganen zu schwimmen. In Gymnospermen werden die Spermien durch Pollenschläuche transportiert. Selbst dann werden Spermien von Cycas und Ginkgo ciliiert.

(ix) Rekapitulationstheorie / biogenetisches Gesetz:
Im Jahr 1828 schlug Von Baer, der Vater der modernen Embryologie, das Gesetz von Baer vor, das besagt, dass während der Embryonalentwicklung die verallgemeinerten Merkmale (wie Gehirn, Rückenmark, Achsenskelett, Aortenbögen usw. allen Wirbeltieren gemeinsam sind) Die besonderen Merkmale (wie nur Haare bei Säugetieren, nur bei Vögeln, nur bei Vierbeinern gefundene Gliedmaßen) unterscheiden die verschiedenen Mitglieder der Gruppe.
Später wurde dieses Gesetz 1866 von Ernst Haeckel als das biogenetische Gesetz modifiziert. Das biogenetische Gesetz von Haeckel besagt, dass "Ontogenese die Phylogenese wiederholt". Ontogenese ist die Lebensgeschichte eines Organismus, während Phylogenie die Evolutionsgeschichte der Rasse dieses Organismus ist. Mit anderen Worten wiederholt ein Organismus seine Ahnengeschichte während seiner Entwicklung.
Beispiele:
(a) In der Entwicklung des Frosches bildet sich eine fischartige Schwanzlarve (Kaulquappe), die mit dem Schwanz schwimmt und an den Kiemen atmet. Dies zeigt an, dass der Frosch aus einem Fisch wie einem Vorfahren entwickelt wurde.
(b) Kaulquappe (Larve) von Herdmania (Urochordat) zeigt Zeichen von Chordaten, dh Vorhandensein von Notochord, gut entwickeltem dorsal platziertem Zentralnervensystem und Schwanz. Die erwachsene Herdmania hat jedoch kein Notochord und keinen Schwanz. Das Nervensystem ist bei erwachsenen Herdmanien ebenfalls stark eingeschränkt. So zeigt die Larve ihre Ahnencharaktere.
(c) Das Protonem, ein frühes Stadium in der Entwicklung eines Mooses und eines Farn-Gametozyten, ähnelt den filamentösen Grünalgen in Struktur, Wachstumsmuster und Physiologie. Dies weist auf eine Algenstamm der Bryophyten und Pteridophyten hin.
(d) Die Gymnospermen sind normalerweise von Wasser bei der Befruchtung unabhängig geworden. Aber die primitiven Gymnospermen (z. B. Cycas und Ginkgo) haben Spermien geißeln und benötigen zur Befruchtung Wasser wie die Pteridophyten. Dies deutet darauf hin, dass die Gymnospermen vom pteridophytenähnlichen Vorfahren abstammen.
4. Biogeographische Nachweise (Nachweise aus der Biogeographie):
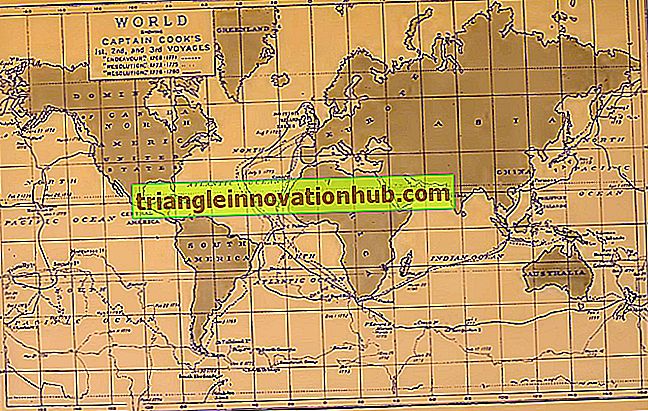
Biogeographie ist die Untersuchung der Verteilung von Tieren und Pflanzen auf dieser Erde. Die Evolutionsbeweise, die auf der Biogeographie (G. Bioslife, Ge Earth, Grapho to Writing) beruhen, werden als biogeographische Beweise bezeichnet. Pangaea (gr. Alle Erde). Es wird vermutet, dass alle Kontinente der heutigen Zeit um die Karbonzeit (vor etwa 345 Millionen Jahren) oder etwas früher in Form einer einzigen großen Landmasse namens Pangaea waren (Abb. 7.37). Später brachen aufgrund verschiedener geologischer Veränderungen riesige Landmassen ab und lösten sich voneinander.
Als sich diese Landmassen (jetzt Kontinente genannt) wegbewegten, wurden sie durch die Meere voneinander getrennt. Die Meere dienten als Barriere und verhinderten die freie Bewegung von Organismen zwischen den Kontinenten. Da diese Kontinente unterschiedliche Umweltbedingungen hatten, entwickelten sich Pflanzen und Tiere unter- schiedlich.
Biogeographische Nachweise können unter den folgenden Überschriften erläutert werden.
1. Biogeographische Bereiche:
Die Erde wurde in sechs große biogeographische Regionen unterteilt, die auf der Grundlage der Verteilung von Tieren und Pflanzen als Bereiche bezeichnet werden. Dr. PL Sclater schlug im Jahre 1858 erstmals die Unterteilung der Welt in sechs Bereiche oder Regionen vor, entsprechend der Verteilung der Vögel. Im Jahr 1876 übernahm AR Wallace es für alle Tiere. Diese Bereiche (Regionen) sind:
(i) Paläarktisches Reich:
Es umfasst Europa, nördlich des Himalaya, China, die Sahara-Wüste Afrikas, Sibirien und einen Großteil Asiens. Wichtige Tiere: Anabas, Bufo, Rhacophorus, Alytes, Proteus, Nekturus, Varanus, Alligator, Falken, Kamel, Tiger, Siegel, Panda.
(ii) orientalischer Bereich:
Es umfasst Indien, Malaysia, die Philippinen, Indonesien, Sri Lanka, Myanmar (Burma). Wichtige Tiere: Karpfen, Katzenfische, Apods, Frösche, Draco, Python, Kobra, Königskobra, Krokodil, Gavialis. Pfau, Hornbills, Stachelschweine, Loris, Gibbon, Nashorn, Elefanten, Tiger, Löwe.
(iii) australischer Bereich:
Es umfasst Australien, Neuseeland und Neuguinea. Wichtige Tiere: Ceratodus (Australischer Lungenfisch), Sphenodon, Casuarius, Emu, Kiwi, Entenschnabeltier, Stachelameisenbär, Opposum, Känguru, Beuteltierkatze.
(iv) Ethopisches Reich:
Es umfasst Afrika, Arabien und Madagaskar. Wichtige Tiere: Protopterus (Afrikanischer Lungenfisch), Rhacophorus, Krokodil, Chamäleon, Python, Strauß, Schuppenfresser, Schimpansen, Gorilla, Zebra, Elefanten, Nilpferd, Nashorn, Giraffe, Löwe, Tiger.
(v) Nearktisches Reich:
Es umfasst Kanada, die Vereinigten Staaten von Amerika und Mexiko. Wichtige Tiere: Saugerfische, Tigersalamander, Amphiuma, Holländer (Giftigeidechse), Alligator, Falke, Opossum, Stachelschwein.

(vi) Neotropischer Bereich:
Es umfasst das Gebiet von Mittel- und Südamerika sowie die Insel Westindiens. Wichtige Tiere: Lepidosiren (südamerikanischer Lungenfisch), Caecilians (Apoda), Hyla, Pipa, Rasselnatter, Rhea, Opossum, Vampirfledermaus, Lama (wie Kamel), Beuteltier Ratte.
Das orientalische Reich ist vom Himalaya-Gebirge vom paläarktischen Reich getrennt. Äthiopisches und australisches Reich sind durch das Meer getrennt.
Orientalisches Reich und australisches Reich werden durch Wallaces Linie getrennt.
Paläarktisches Reich und Nearktisches Reich bilden zusammen eine holoarktische Region.

2. Diskontinuierliche Verbreitung nahe verwandter Arten:
Manchmal treten sehr ähnliche Arten an weit voneinander entfernten Orten auf, ohne dass ein Vertreter in einem Gebiet dazwischen liegt. Dies wird als diskontinuierliche Verteilung bezeichnet. Zwei spezifische Beispiele für diskontinuierliche Verteilung sind unten angegeben.
(a) Alligatoren:
Sie kommen nur im Südosten der USA und im Osten Chinas vor. Der nordamerikanische Kontinent war im frühen Koenozoikum mit Ostasien verbunden. Die Alligatoren waren über die gesamte Region verteilt. Aufgrund bestimmter Barrieren waren die Alligatoren zweier Regionen lange Zeit getrennt und entwickelten einige Mutationen. Daher sind diese Alligatoren etwas unterschiedlich, aber sie sind verwandte Arten derselben Gattung.
(b) Lungenfische:
In frühen Stadien der Kontinentalverschiebung waren Südamerika, Afrika, die Antarktis und Australien miteinander verbunden. Später wurden sie getrennt. Die Antarktis wurde an einen weit entfernten Ort verlegt. Jetzt sind die Lungenfische nur in Südamerika, Afrika und Australien zu finden (Abb. 7.39).

Wenn wir eine Weltkarte auf einem Blatt Papier betrachten, schneiden Sie die Umrisse Südamerikas und Afrikas aus und bringen Sie sie zusammen (Abb. 7.40). Wir finden, dass die rechte Seite Südamerikas in die linke Seite Afrikas passt.

(c) Kamele:
Sie kommen in Asien vor, während ihre nächsten Verbündeten Limas in Südamerika gefunden werden.
(d) Elefanten:
Sie sind in Afrika und Indien zu finden und nicht an Orten mit gleichem Klima in Brasilien.
(e) Tapire:
Sie werden in tropischen Amerika und in malayischen Inseln gefunden.
(f) Magnolien, Tulpen und Sassafras:
Diese Pflanzen wachsen jetzt auf natürliche Weise in den östlichen USA und nur in China. Der Grund ist der gleiche wie für die Alligatoren.
3. Beschränkte Verteilung:
Die vom Festland getrennten Teile haben eine einzigartige Fauna und Flora. Zum Beispiel hat Australien:
(i) Eiablage und
(ii) eingepackte Säugetiere, die nur in Australien vorkommen. Diese eingeschränkte Verteilung kann auf folgende Weise erklärt werden. Australien trennte sich im mesozoischen Zeitalter vom asiatischen Festland, bevor sich Plazentasäugetiere entwickelten. Da die Plazentasäugetiere anpassungsfähiger waren, eliminierten sie die Eiablage und die meisten Säugetiere in anderen Teilen der Welt. Die Eiablage und die beutelten Säugetiere Australiens überlebten, da Plazentagetiere aufgrund fehlender Landroute nicht erreicht werden konnten.
(iii) Wüsten Amerikas besitzen Kakteen, während Afrikas Euphorbien haben,
(iv) Doppelkokosnuss ist auf die Insel Seychles beschränkt.
4. Adaptive Strahlung (= divergente Evolution):
Die Entwicklung unterschiedlicher Funktionsstrukturen aus einer gemeinsamen Ahnenform wird als adaptive Strahlung bezeichnet. Das Konzept der adaptiven Strahlung in der Evolution wurde 1902 von HF Osborn entwickelt. Homologe Organe zeigen adaptive Strahlung.
Beispiele:
(i) Darwin-Finken der Galapagos-Inseln:
Sie hatten gemeinsame Vorfahren, haben jedoch jetzt unterschiedliche Arten von modifizierten Schnäbeln gemäß ihren Ernährungsgewohnheiten (siehe Abbildung 7.46). Darwin unterschied dreizehn Arten von Finken und gruppierte sie in sechs Haupttypen: (a) Große Bodenfinken, (b) Kaktusfinken, die sich von Kakteen ernähren, (c) Vegetarische Baumfinken, (d) Insektenfressende Baumfinken, (e) Warblerfinken (f) Werkzeug- oder Holzpeckfinken.
(ii) australische Beuteltiere:
Darwin erklärte, dass die adaptive Bestrahlung in Australien zu einer Vielzahl von Beuteltieren (gefütterten Säugetieren) geführt habe, und zwar in demselben Prozess der adaptiven Bestrahlung wie in den Finken der Galapagos-Inseln.
(iii) Fortbewegung bei Säugetieren:
Die adaptive Bestrahlung basierend auf der Fortbewegung bei Säugetieren ist ein gutes Beispiel, wie in Abb. 7.42 gezeigt.

5. Konvergente Evolution (= Adaptive Konvergenz):
Die Entwicklung ähnlicher adaptiver Funktionsstrukturen in nicht verwandten Organismengruppen wird als adaptive Konvergenz oder konvergente Evolution bezeichnet.
Beispiele:
(i) Die Flügel von Insekten, Vögeln und Fledermäusen zeigen eine deutliche konvergente Entwicklung.
(ii) Australische Beuteltiere und Plazentasäugetiere zeigen eine konvergente Entwicklung, z. B. den Wolf der Plazenta und das Wolfsmarsupial der Tasmanier.

(iii) Verschiedene nicht eng verwandte Wasserwirbeltiere zeigen eine deutliche konvergente Entwicklung.
(iv) Ameisenbären wie stachelige Ameisenbären und schuppige Ameisenbären gehören zu verschiedenen Klassen von Säugetieren der Klasse, die nicht eng miteinander verwandt sind, aber ähnliche Anpassungen für die Ernährung von Ameisen, Termiten und anderen Insekten erworben haben.
Parallele Entwicklung:
Wenn konvergente Evolution in nahe verwandten Arten gefunden wird, spricht man von „Parallel Evolution“. Beispiel: Entwicklung von Laufgewohnheiten bei Rehen (2-Zehen) und Pferden (1-Zehen) mit zwei Resten von Schienenklingen. Tasmanischer Wolf ist ein Beuteltier, während Wolf ein Plazentasäugetier ist. Dies zeigt auch Parallelität.
5. Nachweise aus der Biochemie und der vergleichenden Physiologie:
Lebewesen weisen eine große Ähnlichkeit in chemischer Konstitution, biochemischen Reaktionen und Körperfunktionen auf. Sie liefern eine Reihe von Beweisen der gemeinsamen Abstammung und der Entwicklung verschiedener Organismengruppen.
1. Protoplasma:
Alle Lebewesen bestehen aus Protoplasma, im Allgemeinen als lebende Materie bezeichnet. Ihre biochemische Konstitution ist in allen Organismen ähnlich. Das Protoplasma besteht zu etwa 90% aus vier Elementen - Kohlenstoff, Wasserstoff, Sauerstoff und Stickstoff. Neben Phosphor und Schwefel bilden sie die meisten organischen Verbindungen der lebenden Materie - Kohlenhydrate, Proteine, Lipide (Fette) und Nukleinsäuren.
2. Nukleinsäuren und Chromosomen:
Das Erbmaterial liegt in Form von DNA vor. DNA ist normalerweise in Chromatinfasern im Kern und in Chromosomen in einer sich teilenden Zelle organisiert. Es hat die gleiche chemische Zusammensetzung in allen Organismen. Genetischer Code, der die Wirkung von DNA-Nukleotiden ausdrückt, ist universell.
3. Enzyme:
Ein Organismus hat mehrere Systeme. Ein System besitzt einen ähnlichen Satz von Enzymen in verschiedenen Organismen, so sehr, dass der Kreb-Zyklus in Pflanzen und Tieren ähnliche Enzyme aufweist. Die Enzyme Trypsin und Amylase sind im gesamten Tierreich gleich. Wirbeltiere haben einen ähnlichen Satz an Verdauungsenzymen in ihrem Verdauungstrakt. Aufgrund dessen können die Verdauungsenzyme eines Tieres sicher einem anderen Tier, einschließlich Menschen, verabreicht werden.
4. Hormone:
Es handelt sich dabei um biochemische Substanzen, die von Drüsen oder endokrinen Drüsen produziert werden, die bei der Auslösung von Reaktionen oder Funktionen in anderen Körperteilen helfen. Die Hormone von Wirbeltieren sind sowohl chemisch als auch funktional ähnlich. Bei einem Mangel an Menschen werden die Hormone anderer Wirbeltiere als Injektionen verabreicht, z. B. Insulin, Thyroxin.
5. Stoffwechsel:
Unterschiedliche Stoffwechselreaktionen wie Atmung, Verdauung, Assimilation, Muskelkontraktion, Nervenleitung (bei Tieren) und Photosynthese (bei Pflanzen) zeigen in verschiedenen Lebewesen eine physiologische Harmonie.
6. Photosynthetische Pigmente:
Alle eukaryotischen autotrophen Pflanzen besitzen Chlorophyll a. Chlorophyll b kommt in Grünalgen und Embryophyten vor. Letztere müssen also aus Grünalgen stammen. Andere Algen besitzen anstelle von b Chlorophyll c, d oder e. Sie müssen von einem gemeinsamen Vorfahren der Algen stammen.
7. Ausscheidung:
Stickstoffhaltige Abfälle zeigen eine progressive Entgiftung bei Wirbeltieren. Es ist Ammoniak bei Fischen, Harnstoff bei Amphibien, Harnsäure bei Reptilien und Vögeln und eine Kombination aus Harnstoff, Harnsäure und anderen Chemikalien bei Säugetieren.
8. Blut und Lymphe:
Blut und Lymphe sind flüssiges Bindegewebe, das bei den meisten Tieren die gleiche Zusammensetzung und Funktion aufweist, was auf eine enge Beziehung hindeutet.
9. Blutgruppen:
Menschen haben vier Hauptblutgruppen - A, B, AB und О. Eine AB-Gruppierung findet sich auch bei Affen, aber nicht bei Affen. Dies zeigt, dass der Mensch mehr mit Affen verwandt ist als mit Affen.
10. Oxyhämoglobinkristalle:
Kristalle, die aus Oxy-Hämoglobin von Wirbeltieren gebildet werden, zeigen eine Beziehung zwischen den Wirbeltieren. Kristalle von nahe verwandten Arten haben das gleiche Muster oder die gleiche Konfiguration, während die von entfernt verwandten Arten eine andere Konfiguration haben. Zum Beispiel haben Vogelkristalle eine grundlegende Ähnlichkeit und zeigen eine grundlegende Ähnlichkeit mit Oxy-Hämoglobinkristallen von Säugetieren, Reptilien und Amphibien.
11. Serumtests (Präzipitin- oder Bluteiweißtests):
Jede Rasse von Individuen hat bestimmte spezifische Proteine, die nicht in anderen Rassen gefunden werden. Eng verwandte Organismen zeigen eine größere Ähnlichkeit dieser spezifischen Proteine als entfernt verwandte Formen. Dies kann durch Präzipitin- oder Serumtests überprüft werden.
Eine Beziehung, die durch Serum-Test zwischen verschiedenen Pflanzengruppen gewonnen wird, ähnelt bekannten evolutionären Tendenzen. In ähnlicher Weise beweisen Blutserumtests verschiedener Wirbeltiergruppen, dass Vögel Krokodilen näher sind als andere Reptilien, während Menschen mit Affen verwandt sind, letzteren mit Affen der alten Welt, Affen der neuen Welt usw.
6. Beweise aus der Zytologie:
Zytologie ist das Studium von Zellen. Es liefert auch Evolutionsbeweise.
(1) Zellularität:
Alle Organismen bestehen aus Zellen und ihren Produkten. Die Zellen können prokaryotisch oder eukaryotisch sein.
(2) Protoplasma:
Alle Zellen bestehen aus einer lebenden Materie namens Protoplasma. Protoplasma wird die physische Basis des Lebens genannt.
(3) Plasmalemma:
Alle Zellen haben eine ähnliche Lipoproteinbedeckung von Plasmalemma oder Plasmamembran oder Zellmembran.
(4) Zellwand:
Es kommt in allen Pflanzenzellen, Pilzen und Bakterienzellen vor.
(5) Kern:
Diese Organelle enthält einen DNA-Proteinkomplex oder Chromatin. Es enthält erbliche Informationen und steuert die Zellaktivitäten.
(6) Ribosomen:
Sie nehmen an der Proteinsynthese teil und werden als "Proteinfabriken" bezeichnet.
(7) Mitochondrien:
Sie sind Sitzplätze für aerobe Atmung und werden als „Krafthäuser“ der Zelle bezeichnet.
(8) Chloroplasten:
Sie führen Photosynthese durch. Ihre Struktur ist in allen Pflanzengruppen im Wesentlichen ähnlich.
(9) Mikroröhrenstrukturen:
Alle Eukaryonten besitzen Mikrotubuli, die Zentriolen, Basalgranulate, Zilien, Flagellen und Spindelapparate bilden.
(10) Zellteilung:
Alle Organismen durchlaufen häufig eine Zellteilung durch Mitose und gelegentlich durch Meiose. Das Muster ist in allen Organismen ähnlich.
7. Taxonomische Nachweise:
Die Organismen, die ähnliche Eigenschaften aufweisen, werden in einer bestimmten Gruppe angeordnet, z. B. werden Fische, Frösche, Eidechsen, Vögel und Menschen als Wirbeltiere zusammengefasst, da alle diese Tiere eine Wirbelsäule besitzen. Die Charaktere einer bestimmten Gruppe müssen entweder im embryonalen Stadium oder im adulten Stadium dieses Organismus vorhanden sein.
Das Tierreich ist in Klassen, Klassen, Ordnungen, Familien, Gattungen, Arten usw. unterteilt. Alle Tiere des Tierreichs sind so in einer Reihe angeordnet, dass die früheren Formen einfacher und die letzteren Formen komplexer sind.
Die zellulären Formen (Protozoen), die am unteren Ende der Evolutionsleiter liegen, sind die einfachsten, während die Säugetiere, die komplexesten Tiere sind, oben liegen. Alle intermediären Tierarten wurden aufgrund ihres unterschiedlichen Entwicklungsstatus zwischen Protozoen und Säugetieren platziert. Diese systematische Anordnung aller Tiere zeigt, dass ein kontinuierlicher Evolutionsprozess stattgefunden hat.
8. Beweise aus der Genetik:
In Organismen treten eine Reihe von Mutationen oder plötzlichen vererbbaren Variationen auf. Sie können in allen Körperteilen und in allen denkbaren Richtungen auftreten. Bei der Akkumulation führen Mutationen zu neuen Arten. Einige wichtige Mutationen sind Ancon Sheep, Double Toed Cats, Hornless Cattle, Rote Sonnenblume, Große Banane usw.
Kaninchen wurden im 15. Jahrhundert auf der Insel Porto Santo eingeführt. Sie haben Mutationen durchgemacht. Heutzutage sind die Kaninchen von Porto Santo kleiner als der Originalbestand, weisen ein anderes Farbmuster auf und sind nächtlicher. Sie züchten nicht mit dem Stammbestand.
Helle Motten Bison betularia unterlag einer Mutation, um aus Biston carbonaria eine Dunkelheit zu erzeugen. Letzteres ist eher für industrielle Bereiche geeignet und hat überlebt, während die weiße Mutterform jetzt auf kleine, unverschmutzte Taschen beschränkt ist.
9. Nachweise aus der Tier- und Tierzucht:
Es gibt Tausende von Sorten Weizen, Zuckerrohr, Reis und andere Kulturpflanzen. In ähnlicher Weise treten Hunderte von Sorten bei Hund, Taube, Pferd und Kuh auf. Büffel, Küken usw. Verschiedene Goldfische stehen zur Verfügung.
Eine derart große Anzahl von Sorten wurde durch Selektion, Hybridisierung, Inzucht und Akkumulation von Mutationen hergestellt. Mutation und Polyploidie werden jetzt künstlich induziert. Neue Spezies wurden durch Allopolyploid (interspezifische Polyploidie) produziert.