Anmerkungen zum Prozess und Mechanismus des Übergangs
In diesem Artikel erfahren Sie mehr über den Prozess und den Mechanismus des Übergangs in der Vererbung!
Janssens (1909) entdeckte als erster die Chiasmenbildung und den damit verbundenen Kreuzungsprozess (Chiasma-Typ-Hypothese). Morgan (1910) fand die Phänomene der Verbindung und Rekombination.

Bild-Höflichkeit: online.santarosa.edu/homepage/cgalt/BIO10-Stuff/Ch09-Genetics/Crossing-Over.jpg
Die Rekombination oder neue Kombination von Genen ist nur durch den Austausch genetischen Materials zwischen homologen Chromosomen möglich (Breakage and Reunion Theory, Darlington 1937). Die Verknüpfung ist in solchen Fällen unvollständig.
Definition:
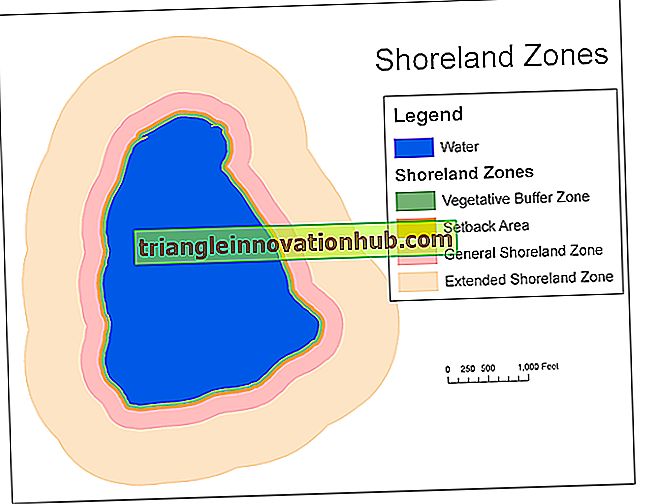
(i) Crossing over ist eine Rekombination von Genen aufgrund des Austauschs von genetischem Material zwischen zwei homologen Chromosomen.
(ii) Es ist der gegenseitige Austausch von Segmenten genetischen Materials zwischen Nicht-Schwesterchromatiden zweier homologer Chromosomen, um Rekombinationen oder neue Kombinationen von Genen zu erzeugen.
Die Nicht-Schwesterchromatiden, bei denen der Austausch von Segmenten stattgefunden hat, werden Rekombinanten oder Cross-Over genannt, während die anderen Chromatiden, bei denen keine Kreuzung stattgefunden hat, als Eltern-Chromatiden oder Nicht-Cross-Over bezeichnet werden.
Beispiel:
CB Hutchinson (1922) kreuzte eine reinrassige farbige und glattkörnige (CS / CS) Maissorte mit reiner Zucht farbloser und geschrumpfter genarbter (cs / cs) Maissorte. Pflanzen der F1-Generation besaßen farbige und glatte Körner, was zeigt, dass beide Ausdrücke dominieren. F 1 -Pflanzen waren offensichtlich heterozygot für die beiden Merkmale (CS / cs). F 1- Pflanzen wurden dann mit doppelt rezessiven Eltern (cs / cs) getestet. Es wurden vier Arten von Nachkommen im Verhältnis 27: 1: 1: 27 anstelle des Verhältnisses 1: 1: 1: 1, das für ein unabhängiges Sortiment erwartet wird (Tabelle 5.3).
Tabelle 5.3. Testkreuz zwischen heterozygot gefärbten und glattkörnigen (CS / cs) Maispflanzen mit doppelt rezessiven Elternteilen mit farblosen und geschrumpften Körnern (cs / cs):
| Art | Häufigkeit von Organismen | Erwartete Häufigkeit bei unabhängigem Sortiment | Häufigkeit erwartet bei vollständiger Kopplung oder Nichtüberfahren |
| Elternarten: 1. Farbig und glatt gemasert | 48, 2% | 25% | 50% |
| 2. Farblos und geschrumpft | 48, 2% | 25% | 50% |
| Rekombinante Typen: 3. Gefärbt und geschrumpft | 1, 8% | 25% | Null |
| 4. Farblos und glatt gemasert | 1, 8% | 25% | Null |
Das obige Verhältnis ist möglich, wenn die Gene der beiden Merkmale kein unabhängiges Sortiment aufweisen. Offensichtlich sind die beiden Gene im selben Chromosom verknüpft, aber 1, 8% Nicht-Schwesterchromatide der homologen Chromosomen zeigen durch Austausch von Segmenten eine Kreuzung zwischen den beiden Genen.
Rekombination und Crossing Over:
Eine neue Gruppierung von Genen oder eine neue Kombination von Zeichen, die sich von den elterlichen Typen unterscheidet, wird Rekombination oder rekombinanter Typ genannt. Es entsteht aufgrund von Überschneidungen, die während der Meiose vor der Gametenbildung auftreten. Die Häufigkeit der Rekombination hängt direkt mit der Häufigkeit des Überkreuzens zusammen.
Manchmal können jedoch zwei oder sogar mehr Überschneidungen gleichzeitig in denselben nicht-Schwesterchromatiden der homologen Chromosomen auftreten, ohne die Häufigkeit der Neukombinationen zu verändern. Daher kann die Häufigkeit des Überkreuzens höher sein als die Häufigkeit der beobachteten Neukombinationen. Die Häufigkeit der Rekombination (Cross-over-Wert, COV) wird anhand der Formel berechnet

Dieser COV von 10, 7% zwischen den Genen des roten Auges und den normalen Flügeln bedeutet, dass diese Gene 10, 7 Einheiten voneinander entfernt auf demselben Chromosom liegen.
Elterliche und rekombinante Typen:
Eine Person enthält zwei homologe Chromosomen jedes Typs. Die zwei Chromosomen werden von den zwei verschiedenen Eltern, Vater und Mutter, erhalten. Daher werden diese Chromosomen auch väterliche und mütterliche Chromosomen genannt. Angenommen, diese beiden Chromosomen besitzen zwei verknüpfte Gene, AB (dominant) in einem und ab (rezessiv) im anderen. Jedes Chromosom besitzt ferner zwei Chromatiden eines ähnlichen Typs (AB, AB; ab, ab).
Wenn zum Zeitpunkt der Gametenbildung oder der Meiose keine Überkreuzung stattfindet, werden nur zwei Arten von Gameten produziert, von denen eine die väterliche (AB) und andere mütterliche (ab) Verbindung trägt. Wenn eine Kreuzung an einer Stelle auftritt, wird ein Chromatid jedes homologen Chromosoms in den Segmentaustausch einbezogen, während das andere Chromatid unverändert bleibt.
So werden vier Arten von Chromatiden nach einer Kreuzung hergestellt - AB, Ab, aB, ab. Zwei davon sind elterlich (AB, ab) und zwei rekombinant (Ab, aB). Nach der Meiose trennen sich die vier Arten von Chromatiden und werden an vier verschiedene Gameten weitergegeben, zwei elterliche und zwei rekombinante (Abb. 5.20).

Wenn der Abstand zwischen zwei verknüpften Genen so ist, dass eine Kreuzung bei jeder Meiose regelmäßig auftritt, produzieren sie zwei elterliche und zwei rekombinante Typen im Verhältnis von 25%: 25%: 25%: 25%. Dies ist ähnlich wie bei einem unabhängigen Sortiment. Mendel hatte Glück, dass drei der sieben Eigenschaften, die er in seinen berühmten Experimenten verwendete, ein regelmäßiges Überkreuzen zeigten.
Daher kann ein unabhängiges Sortiment unter zwei Umständen auftreten:
(a) Gene kommen in verschiedenen nicht-homologen Chromosomen vor
(b) Gene, die im selben Chromosom liegen, aber regelmäßig gekreuzt werden (50% Rekombinanten oder Kreuzungen).
Einflussfaktoren auf Crossing Over (und Verknüpfung):
1. Entfernung:
Der physikalische Abstand zwischen zwei Genen bestimmt sowohl die Stärke der Verknüpfung als auch die Häufigkeit des Übergangs zwischen zwei Genen. Die Stärke der Verknüpfung steigt mit der Nähe der beiden Gene. Auf der anderen Seite steigt die Überkreuzungshäufigkeit mit zunehmender Entfernung zwischen den beiden Genen.
2. Alter:
Die Zunahme des Alters verringert in den meisten Fällen den Grad des Übergangs.
3. Geschlecht:
Männliche Drosophila zeigt wenig Überkreuzung. Das Phänomen des Überkreuzens ist bei der weiblichen Fliege recht häufig. Bei einigen anderen heterogametischen Organismen wird auch bei einem Geschlecht ein vernachlässigbares Überschreiten festgestellt.
4. Röntgenstrahlen:
Die Einwirkung von Röntgenstrahlen erhöht das Auftreten von Überkreuzungen. Whittinghill erzeugte mit Hilfe von Röntgenstrahlen eine Reihe von Überkreuzungen bei männlichen Drosophila.
5. Temperatur:
Temperaturschwankungen erhöhen die Überquerungshäufigkeit.
6. Heterochromatin:
Die Anwesenheit von Zentromeren und heterochromatischen Bereichen (z. B. in der Nähe von Telomeren) verringert die Kreuzungsrate.
7. Chemikalien:
Es wurde festgestellt, dass eine Reihe von Chemikalien, die in den Lebensmitteln enthalten sind, den Grad des Übergangs bei Tieren verändern.
8. Interferenz:
Eine Überkreuzung reduziert das Auftreten einer weiteren Überkreuzung in seiner Nähe. Das Phänomen wird Interferenz genannt. Koinzidenz ist das Verhältnis der beobachteten Doppelkreuzung im Verhältnis zu der erwarteten Doppelkreuzung aufgrund von Nichtinterferenz oder unabhängigem Auftreten. Der Zufall ist gering, wenn die Interferenz hoch ist.
Bedeutung:
1. Überqueren ist ein Mittel, um neue Kombinationen von Genen und damit Merkmalen einzuführen.
2. Es erhöht die Variabilität, was für die natürliche Selektion unter veränderten Bedingungen nützlich ist.
3. Da die Häufigkeit der Kreuzung von der Entfernung zwischen den beiden Genen abhängt, wird dieses Phänomen zur Erstellung von Kopplungs-Chromosomenkarten verwendet.
4. Es hat sich gezeigt, dass Gene im Chromosom linear liegen.
5. Züchter müssen kleine oder große Populationen auswählen, um die erforderlichen Überkreuzungen zu erhalten. Um Kreuzungen zwischen eng verbundenen Genen zu erhalten, ist eine sehr große Population erforderlich.
6. Durch Kreuzung erzeugte nützliche Kombinationen werden von Züchtern aufgenommen, um nützliche neue Sorten von Kulturpflanzen und Tieren zu entwickeln. In Indien wurde durch diese gezielte Auflistung nützlicher Neukombinationen eine grüne Revolution erreicht. Operation Flood oder White Revolution wird auch auf ähnlichen Linien durchgeführt.
Arten der Überquerung:
Crossing over kann einfach, doppelt oder mehrfach sein.
(i) einmaliges Crossing Over:
Die Kreuzung erfolgt an einem Punkt zwischen zwei Nicht-Schwesterchromatiden eines homologen Chromosomenpaares. Es gibt zwei elterliche Typen und zwei Rekombinanten.
(ii) Doppelkreuzung:
In einem homologen Chromosomenpaar tritt die Kreuzung an zwei Stellen auf.
(a) wechselseitige Doppelkreuzung:
Zwei Kreuzungspunkte treten zwischen denselben Nicht-Schwesterchromatiden auf,
(b) komplementäre Überkreuzung:
Die zwei Kreuzungen umfassen drei oder alle vier Chromatiden, so dass die Anzahl der Kreuzungen drei oder vier beträgt
von einem oder keinem elterlichen Typ,
(iii) Mehrfache Kreuzung:
Drei oder mehr Kreuzungspunkte treten im selben homologen Chromosom auf. Doppelkreuzungen und elterliche Typen können auftreten oder auch nicht.
Mechanismus der Überkreuzung:
Chromosomen werden in der S-Phase der Interphase repliziert. Daher sind Leptotenchromosomen doppelsträngig, obwohl die beiden Stränge aufgrund des Vorhandenseins eines Nukleoproteinkomplexes zwischen den Chromatiden nicht sichtbar sind.
(i) Synapsis:
Replizierte, aber scheinbar einzelne homologe Chromosomen liegen nebeneinander mit ähnlichen Genorten der beiden Chromosomen genau gegenüber. Sie tritt im Zygotenen-Stadium der Prophase I auf. Das Phänomen wird Synapsis genannt. Die synaptierten Paare homologer Chromosomen werden Bivalente genannt. Die geringe Menge an nicht repliziertem Chromosom (0, 3%) wird ebenfalls repliziert (Stem und Hotta, 1973). Die zwei homologen Chromosomen werden von einem Synaptinemkomplex zusammengehalten.

(ii) Überqueren:
Es tritt im Pachytenstadium im Vierstrangstadium mit Hilfe von Enzymen (Endonuclease, Exonuclease, R-Protein oder Rekombinase; Stern und Hotta, 1969, 1978) auf. Es kommt zu einem Bruch der Chromatidsegmente, zum Austausch von Nicht-Schwester-Chromatidsegmenten und später zu ihrer Verschmelzung an neuen Orten.
(iii) Tetrad-Stufe:
Der synaptinemale Komplex beginnt sich außer im Bereich der Kreuzung aufzulösen. Daher trennen sich die Chromosomen und die Chromatiden unterscheiden sich an den meisten Stellen. Da ein Bivalent jetzt vier Chromatiden zu haben scheint, wird es als Tetradstadium bezeichnet. Sie tritt im Diplotinstadium der Prophase I auf. Die synaptinemalen Bindungspunkte zwischen den homologen Chromosomen werden als Chiasmen bezeichnet. In späteren Stadien verschieben sich die Chiasmen meist zu den Seiten. Das Phänomen wird als Terminalisierung bezeichnet. Viele von ihnen verschwinden vor der Metaphase I.