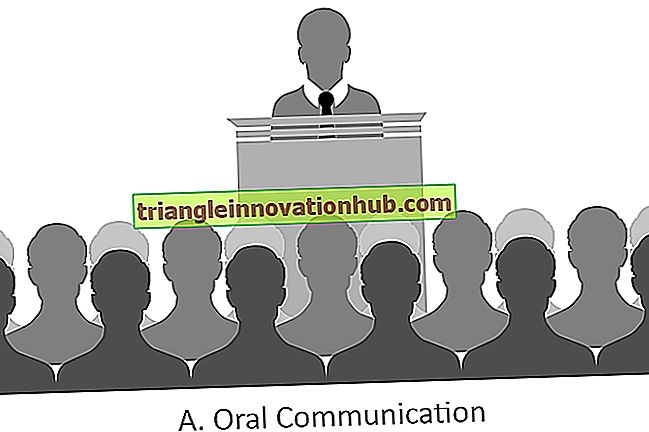
Molekülstruktur der Plasmamembran
Molekülstruktur der Plasmamembran!
Alle biologischen Membranen, einschließlich der Plasmamembran und der inneren Membranen von eukaryotischen Zellen, haben eine gemeinsame Gesamtstruktur: Sie sind Ansammlungen von Lipid- und Proteinmolekülen, die durch nicht-kovalente Wechselwirkungen zusammengehalten werden.
Die Lipidmoleküle sind als kontinuierliche Doppelschicht mit einer Dicke von 4 bis 5 nm angeordnet. Diese Lipiddoppelschicht bildet die Grundstruktur der Membran und dient als relativ undurchlässige Barriere für den Fluss der meisten wasserlöslichen Moleküle.
Die Proteinmoleküle werden in der Lipiddoppelschicht „aufgelöst“ und vermitteln die verschiedenen Funktionen der Membran. einige dienen dazu, spezifische Moleküle in oder in die Zelle zu transportieren; andere sind Enzyme, die Membranreaktionen katalysieren; und wieder andere dienen als strukturelle Verbindungen zwischen dem Zytoskelett der Zelle und der extrazellulären Matrix oder als Rezeptoren zum Empfangen und Transudieren von chemischen Signalen aus der Umgebung der Zelle.
Alle Zellmembranen sind dynamische, fließende Strukturen: Die meisten ihrer Lipid- und Proteinmoleküle können sich schnell in der Membranebene bewegen. Membranen sind auch asymmetrische Strukturen: Die Lipid- und Proteinzusammensetzungen der beiden Flächen unterscheiden sich voneinander in einer Weise, die die unterschiedlichen Funktionen der beiden Oberflächen widerspiegelt.
Obwohl die spezifischen Lipid- und Proteinkomponenten von einem Membrantyp zum anderen stark variieren, sind die meisten grundlegenden strukturellen und funktionellen Konzepte auf intrazelluläre Membranen sowie auf Plasmamembranen anwendbar.
Nachdem wir die Struktur und Organisation der Hauptbestandteile biologischer Membranen (Lipide, Proteine und Kohlenhydrate) betrachtet haben, werden wir die Mechanismen diskutieren, mit denen kleine Moleküle durch ihre Plasmamembranen transportiert werden, und die sehr unterschiedlichen Mechanismen, mit denen sie Makromoleküle und größere Partikel transportieren diese Membran.
Der Lipiddoppelschicht:
Der erste Hinweis darauf, dass die Lipidmoleküle in biologischen Membranen in einer Doppelschicht organisiert sind, stammt aus einem Experiment, das 1925 durchgeführt wurde. Lipide aus roten Blutkörperchenmembranen wurden mit Aceton extrahiert und auf der Wasseroberfläche geschwommen. Die von ihnen besetzte Fläche wurde dann mittels einer beweglichen Barriere verkleinert, bis ein monomolekularer Film (eine Monoschicht) gebildet wurde.
Diese Monoschicht besetzte eine letzte Fläche, die etwa doppelt so groß war wie die ursprüngliche rote Blutzelle, da die einzige Membran in einer roten Blutzelle die Plasmamembran ist. Die Experimentatoren schlussfolgerten, dass die Lipidmoleküle in dieser Membran als durchgehende Doppelschicht angeordnet sein müssen.
Die Schlussfolgerung war richtig, aber es stellte sich heraus, dass sie auf zwei falschen Annahmen beruhte, die sich zufällig kompensierten. Einerseits extrahierte das Aceton nicht das gesamte Lipid. Andererseits beruhte die für die roten Blutkörperchen berechnete Oberfläche auf getrockneten Zubereitungen und war wesentlich geringer als der tatsächliche Wert, der bei Nasszubereitungen zu sehen war.
Daher hatten die aus diesem Experiment gezogenen Schlussfolgerungen einen tiefgreifenden Einfluss auf die Zellbiologie. Infolgedessen wurde die Lipiddoppelschicht zu einem akzeptierten Bestandteil der meisten Membranstrukturmodelle, lange bevor ihre Existenz tatsächlich etabliert war.

Danielli-Davson-Modell oder Protein-Lipid-Protein oder Sandwich-Modell :
Harvey und Cole (1931) wiesen auf die Existenz von Protein durch Untersuchung der Oberflächenspannung von Zellen hin. Dies führte dazu, dass Danielli und Davson ein Lipoproteinmodell der Zellmembran vorschlugen. Gemäß diesem Modell besteht die Plasmamembran aus zwei Schichten von Lipidmolekülen, wie im Lipiddoppelschichtmodell gezeigt.
Die Lipidmoleküle haben ihre Polbereiche an der Außenseite. Man nimmt an, dass Globulin-Proteine mit den polaren Gruppen der Lipide assoziiert sind. Die unpolaren hydrophoben Enden der beiden Lipidschichten liegen einander gegenüber, während ihre polaren hydrophilen Enden durch elektrostatische Wechselwirkung mit Proteinmolekülen verbunden sind. Proteingebundene polare Poren sind in der Membran vorhanden. Diese Poren werden durch periodische Kontinuität der äußeren und inneren Schichten der Plasmamembranproteine gebildet.
Modifikationen des Danielli-Davson-Membranmodells:
Es wurden mehrere Modifikationen der obigen Anordnung beschrieben:
(A) Einige Plasmamembranen haben auf beiden Oberflächen der Lipiddoppelschicht gefaltete ß-Ketten von Proteinen.
(B) Coiled-α-Ketten des helicalen Proteins auf der Oberfläche der Lipiddoppelschicht.
(C) Mit globularen Proteinen auf beiden Oberflächen.
(D) Mit gefalteten Proteinen auf beiden Oberflächen und helikalen Proteinen, die sich in die Poren erstrecken.
(E) Mit gefaltetem ß-Cham-Protein auf einer Seite und globulärem Protein auf der anderen Seite.


Robertsons Einheitsmembranmodell:
Das Einheitsmembranmodell wurde im Jahr 1953 während des Studiums der Zelle unter dem Elektronenmikroskop vorgestellt. Die Grundeinheit der Membranstruktur wurde als allgemein für eine Vielzahl von pflanzlichen und tierischen Zellen angesehen. Alle Zellorganellen wie Golgi-Körper, Mitochondrien, endoplasmatisches Retikulum, Kernmembran usw. haben die Einheitsmembranstruktur.
Die Einheitsmembran wird als trilaminar mit einer bimolekularen Lipidschicht zwischen zwei Proteinschichten betrachtet. Zwei parallele äußere dichte osmiophile Schichten von 20A °, die den beiden Proteinschichten entsprechen. Die mittlere hellfarbige osmiophobe Schicht hat eine Dicke von etwa 35 Å, entsprechend den Kohlenwasserstoffketten der Lipide.
Somit ist die Membraneinheit etwa 7 Å dick. In dieser Hinsicht wird das Danielli-Davson-Modell wieder zusammengefügt. Es unterscheidet sich jedoch vom Danielli-Davson-Modell darin, dass das Protein asymmetrisch ist. An der äußeren Oberfläche befindet sich Mucoprotein, während an der inneren Oberfläche nicht-mukoides Protein vorliegt.

Einwände gegen Einheitsmembran Modell:
Die Einwände gegen das Einheitsmembranmodell nahmen in den 1960er Jahren zu, was zu erneuten Untersuchungen von Lipid-Protein-Wechselwirkungen und zu neuen Modellen führte. Studien von FS Sjostrand (1963) an glatten endoplasmatischen retikulären, mitochondrialen und chloroplastischen Membranen unterstrichen die Unterschiede zwischen den beobachteten Merkmalen von Membranen und die Einheitlichkeit, die für das Einheitsmembrankonzept erforderlich war.

Mitochondrienmembranen und Chloroplastenmembranen enthalten Partikeleinheiten in oder auf der Membran. Die Plasmamembran zeigte nicht das gleiche Aussehen wie Mitochondrien- oder Chloroplastmembranen. Es schien, dass verschiedene Modelle erforderlich sein könnten, um verschiedene Funktionstypen zu beschreiben. Dieser ungeeignete Ansatz wurde unnötig, als ein Mosaikmembranmodell vorgeschlagen wurde.
Größeres Membranmodell:
Wie auch beim Trilaminarmodell liegt auch hier die Lipidschicht zwischen zwei Schichten von Strukturproteinen. Robertson beschrieb die unterschiedlichen Eigenschaften der äußeren und inneren Oberflächen der Membran. Es wurde angenommen, dass die innere Oberfläche mit unkonjugiertem Protein bedeckt ist und die äußere Oberfläche mit Glycoprotein, das den Strukturprotein-Oligosaccharidketten mit negativ geladenen Sialinsäure-Enden überlagert ist, an das Glycoprotein gebunden ist.
Micellar-Modell:
Eine alternative Interpretation der Molekülstruktur der Plasmamembran wurde von Hilleir und Hoffman (1953) postuliert. Sie haben vorgeschlagen, dass biologische Membranen ein nicht-lamellenartiges Muster haben können, das anstelle eines Mosaiks aus globulären Untereinheiten besteht, die als Mizellen bekannt sind, die einen Lipidkern und eine hydrophile Hülle aus polaren Gruppen aufweisen.
Lipidmizellen sind mögliche Bausteine für Membranen, da sie zur spontanen Assoziation neigen. In diesem Modell der Membranstruktur können die Proteinkomponenten der Membran auf beiden Seiten der Ebene der Lipidmicellen eine Monoschicht bilden.
Einzelne Einheiten des Mizellenmosaiks können durch einzelne Enzymmoleküle oder durch Anordnung von Enzymen mit einer präzisen dreidimensionalen Anordnung ersetzt werden, wodurch spezifische Funktionen in die Membranstruktur eingebaut werden können.
Man nimmt an, dass die Räume zwischen den globularen Mizellen wassergefüllte Poren mit einem Durchmesser von 0, 4 nm (4A °) bilden, die teilweise von den polaren Gruppen der Mizellen und teilweise von den polaren Gruppen von assoziierten Proteinmolekülen ausgekleidet sind.

Fluid Mosaic Model:
Dieses Modell wurde von Singer und Nicolson (1972) vorgeschlagen. Gemäß diesem Konzept sind die Lipidmoleküle so angeordnet, dass sie eine eher zusammenhängende Doppelschicht bilden, die den strukturellen Rahmen der Plasmamembran bildet. Die Proteinmoleküle sind auf zwei verschiedene Arten angeordnet. Einige Proteine befinden sich ausschließlich neben den äußeren und inneren Oberflächen der Lipiddoppelschicht und werden als extrinsische Proteine bezeichnet. Andere Proteine dringen teilweise oder vollständig in die Lipiddoppelschicht ein und bilden integrale oder intrinsische Proteine.

Die Lipide und integralen Proteine der Plasmamembran sind von Natur aus amphipatisch. Der Begriff Amphipatie wurde von Hartley (1936) für solche Moleküle geprägt, die sowohl hydrophobe als auch hydrophile Gruppen aufweisen. Die amphipatischen Moleküle neigen dazu, flüssigkristalline Aggregate zu bilden, in denen sich hydrophobe oder unpolare Gruppen innerhalb der Doppelschicht befinden und hydrophile Gruppen auf die Wasserphase gerichtet sind. Daher bilden die Lipidmoleküle eine eher durchgehende Doppelschicht.
Die integralen Proteine sind in der Lipiddoppelschicht interkaliert, wobei ihre Polarregionen von der Oberfläche hervorstehen und nichtpolare Regionen, die in der Lipiddoppelschicht eingebettet sind. Diese Anordnung erklärt, warum die aktiven Zentren von Enzymen und antigenen Glycoproteinen der äußeren Oberfläche der Membranen ausgesetzt sind. Die Quasifluidstruktur der Plasmamembran erklärt die Bewegung der Ansammlung von Proteinmolekülen beträchtlicher Größe über die Membran.
Poren in der Plasmamembran:
Plasmamembran ist durch Poren perforiert. Diese haben einen Durchmesser von etwa 0, 35 nm (Nanometer), etwas größer als die Natriumionen. Weniger als 0, 1 Prozent der Plasmamembran sind durch Poren perforiert, während 99, 9 Prozent der Zelloberfläche für Ionen undurchlässig sind. Es wurden verschiedene Modelle für Porenstrukturen vorgeschlagen.
Einige von ihnen sind:
1. Strukturporen :
Dies sind permanente zylindrische Löcher, die die ansonsten durchgehende Doppelschicht unterbrechen.
2. Dynamische Poren:
Diese Poren sind eher durchgehende zylindrische Löcher als dauerhaft. Sie erscheinen nur zum Zeitpunkt der Einnahme.
3. Pflasterkanalporen :
Gemäß diesem Konzept werden die Poren als die Ecken der dicht gefüllten, fast sechseckigen Pflasterblöcke der Lipid- und Proteinuntereinheiten angesehen.
4. Proteinkanalporen :
Diese Poren werden als Teile des Lipid-Globular-Protein-Mosaikmodells betrachtet. Diese bilden kleine Kanäle aus spezifischen Proteinen, die in die Membran eingebettet sind und durch die Ionen und kleine Moleküle diffundieren können.
5. Ionophor:
Die Ionophore sind kleine Polypeptide, deren eines Ende hydrophob und das andere hydrophil ist. Das hydrophobe (äußere) Ende löst sich in der Membran auf, während das hydrophile Ende (innere Seite) Ionen oder wasserlösliche Materialien aufnimmt und sie auf der anderen Seite ablegt. Die Ionophore helfen beim Austausch von Substanzen aus oder in die Zelle.
Spezialisierung oder Modifikationen :
Mit der erhöhten Auflösung des Elektronenmikroskops wurde eine zahlreiche Spezialisierung der Zelloberfläche erkannt (Sjostrand 1956; Fawcett 1958). Die folgende Beschreibung von Fawcett (1958) befasst sich mit den verschiedenen topografisch untersuchten Spezialisierungen von Plasmamembranen.
Microvilli:
Im Darmepithel sind Mikrovilli sehr prominent und bilden eine kompakte Struktur, die unter dem Lichtmikroskop als gestreifte Grenze erscheint. Diese Mikrovilli, die 0, 6 bis 0, 8 um lang sind und einen Durchmesser von 0, 1 um haben, repräsentieren zytoplasmatische Prozesse, die von der Plasmamembran bedeckt werden. Innerhalb des Zytoplasmakerns werden feine Mikrofilamente beobachtet, die im Zytoplasma ein terminales Netz bilden.
Die Oberfläche der Mikrovilli ist mit einer Schicht aus filamentösem Material (Fuzzy-Mantel) bedeckt, die aus Glykoprotein-Makromolekülen besteht. Microvilli erhöhen die effektive Absorptionsfläche. Zum Beispiel kann eine einzelne Zelle bis zu 300 Mikrovilli haben, und in einem Quadratmillimeter Darm können 200, 0 und 000 vorhanden sein. Die engen Räume zwischen den Mikrovilli bilden eine Art Sieb, durch das Substanzen während der Absorption passieren müssen.
Zahlreiche andere Zellen haben neben Darmepithel Mikrovilli, wenn auch weniger. Sie wurden in Mesothelzellen, in den Epithelzellen der Gallenblase, der Gebärmutter und des Dottersacks, in Leberzellen usw. gefunden.
Die Bürstenumrandung des Nierentubulus ist der gestreiften Umrandung ähnlich, obwohl sie größere Abmessungen aufweist. Eine amorphe Substanz zwischen den Mikrovilli gibt eine periodische Säure-Schiff-Reaktion für Polysaccharide an. Zwischen den Mikrovilli, an der Basis, dringt die Zellmembran in das apikale Zytoplasma ein. Bei diesen Invaginationen handelt es sich offenbar um Wege, über die große Mengen an Strömung durch einen der Pinocytose ähnlichen Vorgang eintreten.

Desmosomen oder Macula Adherens:
Desmosomen sind Zellverbindungen, die hauptsächlich in Zellen eines einfachen säulenförmigen Epithels vorkommen. Diese treten als spezialisierte Bereiche entlang der Kontaktflächen auf. Unter dem Lichtmikroskop werden die Desmosomen als dunkel gefärbte Körper gesehen. Unter dem Elektronenmikroskop erscheinen diese als knopfförmige Verdickungen an der Innenfläche von Plasmamembranen benachbarter Zellen am Kontaktpunkt.
Die Verdickungen werden von feinen zytoplasmatischen Fibrillen, den sogenannten Tonfibrillen, durchquert, die im weiten Bogen eine Art Schleife bilden. Diese Filamente stabilisieren die Verbindung und dienen als Verankerungsstellen für die zytoplasmatischen Strukturen. Die Plasmamembranen benachbarter Zellen in den Bereichen von Desmosomen sind durch einen Interzellularraum von etwa 30 bis 35 nm getrennt. Es ist mit dazwischenliegendem dichtem Beschichtungsmaterial gefüllt, das in der Mitte eine dunkle Linie bildet. Es besteht aus Mucopolysacchariden und Proteinen.
Die Desmosomen befassen sich in erster Linie mit der Zelladhäsion, helfen aber auch dabei, die Zellform aufrechtzuerhalten, indem sie Starrheit und Zellunterstützung bieten. Ersteres wird durch die interzelluläre Beschichtungssubstanz und letztere durch die Tonofibrillen bewirkt.

Plasmodesmen :
Manchmal sind die Zellen durch Brücken des Cytoplasmas verbunden, die zwischen den Poren der Zellwand oder der Plasmamembran zwischen den benachbarten Zellen durchlaufen. Solche Verbindungen werden Plasmodesmen genannt. Sie sind normalerweise einfach, aber es kann auch ein Astomosierendes Plasmodesmus gefunden werden. Ihre Verteilung und Anzahl kann auch sehr erheblich sein. Sie wurden von Tang entdeckt! (1879) und wurden von Strasburger (1882) so benannt.
Endoplasmatisches Retikulum ist an den Stellen, an denen Plasmodesmen vorhanden sind, oft eng mit der Zelloberfläche verbunden. Durch sie wird oft die zytoplasmatische Kontinuität zwischen den benachbarten Zellen aufrechterhalten. Sie bieten einen Mittelwert für die Interaktion zwischen benachbarten Zellen, die in anderen Regionen getrennt sind. Durch sie kann das Material von Zelle zu Zelle gelangen.
Es ist nicht bekannt, ob alle Plasmodesmen einander ähnlich sind. Es gibt einige Unterschiede, da sie nicht nur zu der Zeit produziert werden, wenn sich die Zellen teilen, sondern sich auch spontan zwischen Zellen gebildet haben, die miteinander in Kontakt gekommen sind, z. B. Tylosen in Xylem-Gefäßelementen.
Sie können einzeln auftreten oder zu Gruppen zusammengefasst werden. In vielen Primärwänden sind die Plasmodesmen normalerweise mit einer reduzierten Anordnung von Wandmaterial verbunden, und die Fläche wird dann als Primärgrube oder -feld bezeichnet.

Hemi-Desmosomen:
Diese befinden sich in der Grundfläche einiger Epithelzellen. Ihre Struktur ist den Desmosomen ähnlich, aber diese werden durch eine Hälfte dargestellt; Ihr Gegenstück wird normalerweise durch Collegenfibrillen dargestellt.
Terminalleisten :
Die Anschlussstäbe werden auch als Zwischenverbindungen oder Zonula Adhaerens bezeichnet. Die terminalen Stäbe sind den Desmonomen ähnlich, außer dass ihnen die Tonofibrillen fehlen. Im Endriegel ist die Plasmamembran verdickt und das Cytoplasma der verdickten Fläche ist dicht. Die terminalen Stäbe kommen im Zwischenteil der Plasmamembran von säulenförmigen Zellen unmittelbar unter der Oberfläche vor. Die korrekte Identität von Zonula Adhaerens ist immer noch fraglich (JP Trinkaus, 1969).
Membran-Wechselwirkungen:
Ein weiterer Aspekt von Zellmembranen, der eine Diskussion verdient, ist die Interaktion zwischen Membranen verschiedener Zellen. Interzelluläre Kommunikation ist für viele Zellfunktionen und insbesondere während der Entwicklung des Organismus wichtig, wenn Zellen ständig mit anderen Zellen interagieren.
Die Art der Membranwechselwirkungen kann von vollständigen zytoplasmatischen Brücken zwischen Zellen bis zu lokalisierten Bereichen von Membranverbindungen variieren, die einen Kontaktbereich von nur wenigen Angström oder bis zu mehreren Mikrometern umfassen können. Der strukturelle Charakter des tatsächlichen Kontakts fällt im Allgemeinen in eine von drei Kategorien. Gap Junctions, Tight Junctions und septierte Junctions.

Spaltübergänge erscheinen bei Betrachtung mit dem Elektronenmikroskop als mehrschichtige Strukturen. Sie scheinen zwei Membranen zu sein, die dicht beieinander liegen und einen Spalt von 20 bis 40 Å aufweisen. Die Gesamtdicke des gesamten Spaltübergangs beträgt 170 bis 190 A ° und wird sowohl bei Wirbeltieren als auch bei Wirbellosen gefunden. Sie werden nicht in Skelettmuskelfasern oder roten Blutkörperchen gefunden.

Enge Verbindungen werden nur in Wirbeltieren gefunden und kommen in Zellen wie Epithelzellen vor. Diese Verbindungen scheinen echte Fusionen zwischen der ® Membran zu sein und sie sind 100 bis 140 Å dick.
Septate-Übergänge wurden nur bei wirbellosen Tieren gefunden. Sie sind viel größer als die anderen Arten von Kontakten und zeichnen sich durch elektronendichte Kreuzbrücken aus, die sich zwischen den beiden Zellmembranen erstrecken.