Golgi-Komplex: Terminologie, Vorkommen, Morphologie, Struktur und andere Details
Golgi-Komplex: Terminologie, Vorkommen, Morphologie, Struktur, strukturelle und biochemische Polarisation, chemische Zusammensetzung, Funktionen und Ursprung!
1898 entdeckte Golgi mittels einer Silberfärbungsmethode eine retikuläre Struktur im Zytoplasma. Der Name "Golgi-Apparat", der dieser Struktur im Allgemeinen gegeben wird, ist verwirrend, da er auf eine bestimmte Beziehung zu den physiologischen Prozessen der Zelle hinweist.
Heutzutage scheint es sinnvoller, den Namen „Golgi-Substanz“ oder „Golgi-Komplex“ zu verwenden, um auf dieses Material zu verweisen, das besondere Färbungseigenschaften aufweist. Da der Brechungsindex dem der Matrix ähnlich ist, ist der Golgi-Komplex in lebenden Zellen schwer zu beobachten. Die Verwendung des Elektronenmikroskops hat zu einem deutlichen Bild dieser Komponente geführt, und ihre submikroskopische Struktur wurde gezeigt.
Der Golgi-Komplex galt jahrelang als Artefakt verschiedener Fixierungs- und Färbeverfahren. Mit anderen Worten, viele Wissenschaftler glaubten, dass die Struktur, die während zahlreicher Mikroskopieverfahren beobachtet und als Golgi bezeichnet wurde, in der lebenden Zelle nicht wirklich existierte. Guilliermond (1923), Parat (1927), Walker und A lien (1921), Zweifel an der Existenz des Golgi-Komplexes.
Terminologie des Golgi-Komplexes:
Holmgren bezeichnete den Golgi-Komplex als Trophospongium (Cajal als Golgi-Holmgren-Kanäle). Baker verwendete den Begriff Lipochondrie wegen des vermuteten Lipidgehalts. Der Begriff Dalton Complex wurde 1952 nach dem Namen seines Beobachters Dalton benannt. Sjostrand schlug den Begriff Cytomembranen für das Golgi-System vor.
Sosa hat die folgende Nomenklatur für den Golgi-Komplex vorgeschlagen:
1. Golgiokinese :
Division des Golgi-Apparats während der Kernteilung.
2. Golgiosomes :
Durch die Golgiogenese erzeugte Corpuskel werden als Golgiosomen bezeichnet, die bei Wirbellosen als Golgi-Material bezeichnet werden.
3. Golgiolyse :
Auflösungsprozess des Golgi-Apparats.
4. Gogiorrhexis :
Fragmentierung am Golgi-Apparat.
5. Golegiogenese :
Bildung und Differenzierung des Golgi-Körpers während der Embryonalentwicklung.
6. Golgio-cytoarchitecture :
Untersuchung der Zellstruktur in Bezug auf den Golgi-Apparat.
Auftreten:
Der Golgi-Komplex kommt in allen Zellen außer den prokaryotischen Zellen (Mycoplasma, Bakterien und Blaualgen) und eukaryotischen Zellen bestimmter Pilze, Samenzellen von Bryophyten und Pteridophyten, Zellen von reifen Siebtuben von Pflanzen und reifen Spermien und rotem Blut vor Zellen von Tieren.
Morphologie des Golgi-Komplexes:
Die Morphologie des Golgi-Komplexes variiert von Zelle zu Zelle, je nachdem, um welchen Zelltyp es sich handelt. Zwei Formen des Golgi-Komplexes wurden beobachtet.

1. Lokalisiertes Formular :
In polarisierten Zellen von Wirbeltieren (die eine Basis und einen Scheitelpunkt haben) tritt der Golgi-Komplex einzeln auf und nimmt eine feste Position ein. Es liegt zwischen dem Kern und dem Sekretor. Dies kann am besten in Schilddrüsenzellen, in exokrinen Zellen der Bauchspeicheldrüse und den Schleimzellen des Darms gesehen werden.
2. Diffuse Form :
In einigen spezialisierten Zellen von Wirbeltieren (Nervenzellen und Leberzellen), in den meisten Pflanzenzellen und in den Zellen von Invertebraten werden mehrere Einheiten des Golgi-Komplexes zusammen mit den Elementen des endoplasmatischen Retikulums gefunden. Jede Einheit wird Dictyosom genannt. In Leberzellen kommen bis zu 50 Dictyosomen pro Zelle vor und in bestimmten Pflanzenzellen kann deren Anzahl bis zu Hunderten betragen.
Form :
Die Form des Golgi-Komplexes ist bei verschiedenen somatischen Zelltypen von Tieren ziemlich variabel. Selbst in derselben Zelle gibt es Variationen in verschiedenen Funktionsstadien. Die Form ist jedoch bei jedem Zelltyp konstant. Die Form variiert von einer kompakten Masse bis zu einem dispersen Filamentnetzwerk.
Nummer:
Die Anzahl der Golgi-Stapel pro Zelle variiert je nach Zelltyp enorm - von nur einem bis zu Hunderten. In einigen Zellen gibt es ein einzelnes großes, während es bei Paramoeba zwei gibt. In Stereomyxa (einer Amöbenart) gibt es viele Golgi-Komplexe. Nervenzellen, Leberzellen und die meisten Pflanzenzellen haben auch mehrere Golgi-Komplexe, von denen etwa 50 in Leberzellen vorkommen.
In Algen-Rhizoiden sieht man mehr als 25.000 Golgi-Komplex (Sievers 1965). In einigen spezialisierten Zellen kann der Golgi-Komplex sogar einen großen Teil des Zellvolumens ausmachen. Ein Beispiel ist die Becherzelle des Darmepithels, die Schleim in den Darm abgibt; Die Glykoproteine im Schleim sind hauptsächlich im Golgi-Komplex glykosyliert.
Größe:
Die Größe ist ebenfalls variabel. Es ist groß in den Nerven- und Drüsenzellen und klein in den Muskelzellen. Im Allgemeinen ist der Golgi-Komplex gut entwickelt, während sich die Zelle im aktiven Zustand befindet. Wenn die Zelle alt wird, nimmt der Komplex allmählich ab und verschwindet.
Position:
Die Position des Golgi-Komplexes ist für jeden Zelltyp relativ festgelegt. In den Zellen des ektodermalen Ursprungs ist der Golgi-Komplex ab dem Zeitpunkt des Embryonalzustands zwischen Kern und Peripherie polarisiert (Cajal, 1914). In den sekretorischen exokrinen Zellen, die im Allgemeinen eine typische Polarisation aufweisen, befindet sich der Golgi-Komplex zwischen dem Kern und dem sekretorischen Pol.
In den endokrinen Drüsen ist die Polarität dieses Organoids variabel, außer in der Schilddrüse, wo es auf das Zentrum des Follikels ausgerichtet ist. In den jüngeren Zellen und oft in den älteren liegt sie meistens auf einer Seite des Zellkerns, kann sie jedoch in bestimmten Fällen vollständig umgeben. In Ganglionazellen der Maus ist die Position perinuklear.
Detaillierte Struktur des Golgi-Komplexes:
Dalton und Felix (1954) beschrieben den Golgi-Komplex in den Nebenhoden der Ratte nach der ersten elektronenmikroskopischen Aufnahme.
Die folgende Beschreibung des Golgi-Komplexes basiert auf der Arbeit mehrerer Autoren:
1. Zisternen :
Die Zisternen oder Saccules ähneln der glatten Oberfläche ER und erscheinen im Schnitt als Stapel eng beieinander liegender, von Membranen begrenzter Beutel. Die Anzahl der Saccules variiert bei den meisten tierischen und pflanzlichen Zelltypen zwischen 4 und 8. In Euglena kann die Anzahl auf 20 steigen.
Die Membran der Saccules hat eine Dicke von etwa 60 bis 70 Å, die einen etwa 150 Å breiten Hohlraum umschließt, dessen Ränder oft erweitert sind. Den meisten Autoren zufolge gibt es zwei klar definierte Gesichter der Zisternen, dh konvex und konkav. Letztere wird im Allgemeinen als die reife oder formende oder distale Fläche bezeichnet, und die konvexe Seite wird als unreife oder erregte oder proximale Fläche betrachtet. Die in einer parallelen Anordnung befindlichen Cisterane sind durch einen Abstand von etwa 200 bis 300 A voneinander getrennt 0
Was sie zusammenhält, ist noch nicht bekannt, aber in wenigen Zellen ist zwischen den Saccules eine dünne Schicht aus elektronenundurchlässigem, manchmal dichtem Material zu sehen, die in bestimmten Regionen stärker von Bedeutung ist, auf die Amos und Grimstone (1968) die Begriffsknoten angewandt haben. Mollenhauer et al. (1973) untersuchten in gewissen Abschnitten Intercisternal-Elemente und Plaques in bestimmten Pflanzen-Golgi-Komplexen.

2. Röhrchen :
Aus dem Randbereich der Zisternen entsteht ein komplexes, anastomosierendes flaches Netz von Tubuli mit einem Durchmesser von 300 bis 500 A °. Clowes und Juniper (1969) haben dieses Röhrennetz mit Bandscheiben verglichen.
3. Vesikel :
Die Vesikel sind kleine tröpfchenartige Säcke, die an den Tubuli am Rand der Zisternen haften bleiben.
Sie haben folgende zwei Arten:
(a) glatte Vesikel :
Die glatten Vesikel haben einen Durchmesser von 20 bis 80 µm. Sie enthalten sekretorisches Material (werden oft als sekretorische Vesikel bezeichnet) und werden von den Enden der Zistelkanälchen im Netz abgehoben. Oft sind es mehr als eine Tubuli-Verbindung, und vermutlich füllen sie ein einzelnes sich bildendes Vesikel.
(b) beschichtete Vesikel :
Die beschichteten Vesikel sind kugelförmige Ausstülpungen mit einem Durchmesser von etwa 50 um und einer rauen Oberfläche. Sie befinden sich an der Peripherie der Organelle, gewöhnlich an den Enden einzelner Tubuli, und unterscheiden sich morphologisch von den sekretorischen Vesikeln. Ihre Funktion ist unbekannt.
4. Golgianische Vakuolen :
Dies sind große, runde Säcke, die sich auf dem ausgereiften Gesicht von Golgi befinden. Diese werden entweder durch die expandierten Zisternen oder durch die Fusion sekretorischer Vesikel gebildet. Die Vakuolen sind mit etwas amorpher oder körniger Substanz gefüllt.
Der Golgi-Komplex ist strukturell und biochemisch polarisiert:
Der Golgi-Komplex hat zwei verschiedene Gesichter: ein Cis-oder Formiergesicht und ein Trans- oder Reifegesicht. Die cis-Fläche ist eng mit einem glatten Übergangsbereich des groben ER verbunden. In sekretorischen Zellen ist die trans-Fläche die der Plasmamembran am nächsten liegende Seite: Hier werden die großen Sekretvesikel ausschließlich in Verbindung mit der trans-Fläche eines Golgi-Stapels gefunden, und die Membran eines sich bildenden Sekretvesikels ist oft zusammenhängend mit der von die transseite der letzten ("trans-most") cisterna.
Im Gegensatz dazu sind die kleinen Golgi-Vesikel entlang des Stapels gleichmäßiger lokalisiert, es wird allgemein angenommen, dass Proteine aus dem ER auf der cis-Seite in einen Golgi-Stapel eintreten und für mehrere Ziele auf der Trans-Seite austreten. Es ist jedoch weder der genaue Weg durch den Golgi-Komplex bekannt, noch wie sie entlang der einzelnen Stapel von Cisterna zu Cisterna reisen.
Die zwei Gesichter des Golgi-Komplexes sind biochemisch verschieden. Zum Beispiel kann in bestimmten Fällen eine Variation der Dicke der Golgi-Membranen über den Stapel hinweg detektiert werden, wobei die an der cis-Seite dünner (ER-artig) und die an der trans-Seite dicker sind (plasmamembranartig).
Auffälliger sind die Ergebnisse, die erzielt werden, wenn bestimmte histochemische Tests in Verbindung mit der Elektronenmikroskopie verwendet werden, um bestimmte Proteine innerhalb des Golgi-Komplexes zu lokalisieren. Einige dieser Tests zeigen membrangebundene Enzyme, Aktivitäten, die eine eindeutige Polarität in ihrer Lokalisation innerhalb des Golgi-Stacks aufweisen.
Ein besonders faszinierender biochemischer Befund war die Entdeckung, dass lysosomale Enzyme, wie saure Phosphatase, mit den trans-most-Cisterna des Golgi-Stacks und in einigen der beschichteten Vesikel in der Nähe konzentriert sind. Dies legt nahe, dass spezifische Vesikel, die Lysosomen verlassen, sich in dieser Region befinden.
Sekretorische Proteine werden durch histochemische Verfahren in allen gestapelten Zisternen gefunden, obwohl die großen Sekretvesikel, in denen diese Produkte konzentriert sind, nur mit der Golgi-Cisterna trans verbunden sind.
Chemische Zusammensetzung:
Bezüglich der chemischen Zusammensetzung des Golgi-Komplexes wurde nachgewiesen, dass die folgenden Substanzen vorhanden sind:
1. Phospholipide :
Die Zusammensetzung der Phospholipide von Golgi-Membranen liegt zwischen denen der endoplasmatischen Membranen und der Plasmamembranen.
2. Proteine und Enzyme :
Der Golgi-Komplex aus verschiedenen pflanzlichen und tierischen Zellen weist große Unterschiede im Protein- und Enzymgehalt auf. Einige der Enzyme sind ADPase, ATPase, NADPH-Cytochrom-C-Reduktase, Glykosyltransferasen, Galactosyltransferase, Thiaminpyrophosphat usw.
3. Kohlenhydrate :
Sowohl pflanzliche als auch tierische Zellen weisen einige übliche Kohlenhydratkomponenten auf, wie Glucosarin, Galactose, Glucose, Mannose und Fructose. Pflanzengolgi fehlt Sialinsäure, sie kommt jedoch in hoher Konzentration in der Rattenleber vor. Einige Kohlenhydrate wie Xylulose und Arabinose sind nur in Pflanzenzellen vorhanden.
4. Vitamin С :
Der im Golgi-Komplex gespeicherte Anteil an Vitamin C wurde von Tomitte gezeigt. Ihm zufolge speichert der Golgi-Komplex das Vitamin С und setzt es langsam in ausreichender Menge im Zytoplasma frei, um eine Oxidation der Zellprodukte zu verhindern.

Funktionen des Golgi-Komplexes:
1. Bildung des Akrosoms während der Spermiogenese :
Während der Reifung von Spermien spielt der Golgi-Komplex eine Rolle bei der Bildung von Akrosomen (Burgos und Fawcett, 1955).

In frühen Stadien erscheint der Golgi als kugelförmiger Körper, der aus Zisternen besteht, die in parallelen Stapeln angeordnet sind, und zahlreiche kleine Bläschen. Letzterer wurde immer von den Zisternen abgeklemmt. Mit fortschreitender Entwicklung nimmt der Golgi-Komplex eine unregelmäßige Form an und große Vaculolen werden durch Dilatationen der Zisternenbeutel gebildet.
In der Mitte dieser großen Vakuolen oder Vakuolen befindet sich ein dichtes Granulat, das proacrosomale Granulat. Dieses Granulat, das aus dem Golgi-Komplex stammt, wächst in der Vakuole durch einen als Akkretion bekannten Prozess weiter. Diese Vakuole und dieses Granulat nähert sich dem vorderen Pol der Kernmembran und bildet ein akrosomales Granulat.
Mit der Verlängerung der Spermatide breitet sich das akrosomale Vesikel über die Kernoberfläche aus und bricht schließlich mit der Kernmembran zusammen, wodurch das Kappenmaterial gebildet wird. Das akrosomale Granulat wird zum Akrosom, das an der Spitze des Kerns liegt und offenbar bestimmte Enzyme umfasst, die an dem Befruchtungsprozess beteiligt sind.
2. Synthese und Sekretion von Polysacchariden:
Studien an Becherzellen durch Autoradiographie und Elektronenmikroskopie haben die Wechselbeziehung zwischen Proteinsynthese, Kohlenhydratzugabe und Sulfatierung nachgewiesen. Die Becherzellen des Kolons produzieren Mucigen. Dieses Sekretionsmaterial enthält einen großen Anteil an Kohlenhydraten.
Der Golgi-Komplex befindet sich direkt über dem Kern. In Richtung der freien Oberfläche der Zelle werden die Mucigen-Granulate allmählich vergrößert. Die proximalen Zisternen des Golgi-Komplexes zeigen keine Schwellung, aber in einiger Entfernung über den Stapel werden die distalen Zisternen ziemlich plötzlich in mucigen Granulat umgewandelt. Die distalen Zisternen wandeln sich alle 2-4 Minuten kontinuierlich in mucigen Granulat um. Zum Ausgleich werden neue proximale Zisternen gebildet.
3. Rolle in der Sekretion:
Es wird angenommen, dass der Golgi-Komplex eine Rolle in der Sekretionsfunktion einer Zelle spielt. Die Frage ist jedoch, dass sie einige Substanzen selbst sekretieren oder synthetisieren oder es handelt sich lediglich um ein Lagerhaus, in dem die Sekretionsprodukte, die anderswo in der Zelle ausgeschieden werden, einfach gelagert und konzentriert werden.


Aus den Studien von Palade et al. 1962 ist dieser Sekretionszyklus jetzt genau definiert und umfasst vier Schritte für Pankreas-Azinuszellen.
(i) Der Einbau von Aminosäuren in Protein an der Oberfläche des rauen endoplasmatischen Retikulums.
(ii) Transfer dieser entstehenden sekretorischen Proteine in die Zisternen des rauen endoplasmatischen Retikulums.
(iii) Der intrazelluläre Transport dieser Proteine zum Golgi-Komplex.
(iv) Die Wanderung von Zymogengranulaten zum Scheitelpunkt der Zelle, wo sie in Lumen abgegeben werden.
4. Rolle des Golgi-Körpers in der Oogenese :
Srivastava (1965) hat während der Oogenese einen kurzen Überblick über den Golgi-Komplex gegeben. Nach Afzelius (1956) besteht der Golgi-Komplex eines Seeigel-Eies unter Elektronenmikroskopie aus Lamellenstapeln, die Wände von flachen Beuteln bilden, die gelegentlich geschwollen sein können.
Es gibt einige Hinweise auf Querteilung dieser Körper. Sotelo (1959) und Sotelo und Porter (1959) haben den Golgi-Komplex in Ratten-Eizellen unter dem Elektronenmikroskop beschrieben und in den frühen Oozyten eine juxtra-Kernlokalisierung dieser Organelle gefunden.
Im nächsten Stadium lösen sich diese in Fragmente auf und im dritten Stadium bewegen sie sich in Richtung der Kortikalis. In all diesen Fällen bleibt ihre Struktur eng gepackt aus schlanken Doppelprofilen (abgeflachten Säcken) und sphärischen Vesikeln.
In den frühen Eizellen ist der Komplex kompakt organisiert. In späteren Stadien werden diskrete Bündel von Profilen, die von kleinen Vesikeln umgeben sind, in der kortikalen Zone gefunden. In den frühen Eizellen sind der Golgi-Komplex und das Zentrosom eng miteinander verbunden.
5. Absorption von Verbindungen :
Hirsch et al. Haben entdeckt, dass bei der Verfütterung von Eisenzucker an ein Tier Eisen am Golgi-Komplex (Kedrowsky) absorbiert wird. Van Teel hat gezeigt, dass Golgi-Systeme auch Verbindungen von Kupfer und Gold absorbieren. Kedrowsky hat gezeigt, dass der Golgi-Komplex von Opalina Bismutose (Albumin-Bismut-Verbindung) und Protargol (Albumin-Silber-Verbindung) aufnehmen kann. So geben Kirkman und Severinghaus an, dass der Golgi-Komplex als Kondensationsmembran für die Konzentration von Produkten in Tröpfchen oder Granulat wirkt.
6. Pflanzenzellwandbildung :
Die Zellwand von Pflanzen besteht aus Fibrillen, die hauptsächlich Polysaccharide enthalten, sowie einige Lipide und Proteine. Während der Zytokinese bildet sich zwischen den beiden Tochterkernen eine Zellplatte, um die sich eine Membran befindet, die später zur Plasmamembran der Tochterzellen wird. Es gibt klare Belege dafür, dass die Polysaccharide im Golgi-Komplex gebildet und in die neue Zellwand überführt werden, die während des Wachstums der Zellen abgelegt wird.

Substanzen wie Pektine und Hemicellulosen, die die Matrix der die Plasmamembranen trennenden Zellplatte bilden, werden auch vom Golgi-Komplex beigesteuert.
7. Bildung intrazellulärer Kristalle :
In der Meeresisopode, Limnoria lingmorum, die eine Grabform darstellt, gibt es Mittelgebirge, deren Zellen aus Kristallen bestehen. Diese haben eine Länge von bis zu 30 A ° und eine Dicke von 15 A °. Es wurde nachgewiesen, dass diese Kristalle vom Golgi-Komplex gebildet werden und bekanntermaßen Eiweiß und Eisen enthalten. Sie sind ohne umschließende Membran und meist kugelförmig. Sie befassen sich mit der Sekretionsaktivität.
8. Milcheiweißtröpfchenbildung :
In der laktierenden Brustdrüse von Mäusen werden Proteintröpfchen produziert, die mit dem Golgi-Komplex verwandt sind. Diese Tröpfchen öffnen sich normalerweise auf der Zelloberfläche durch die Verschmelzung ihrer einschließenden Membran mit der Plasmamembran.
9. Bildung von Lysosomen und Vakuolen :
Primäre Lysosomen werden auf dieselbe Weise aus den Golgi-Membranen gebildet wie die sekretorischen Vesikel. Es gibt gute Beweise dafür, dass Dictyosomen in ihren reiferen Regionen hydrolytische Enzyme akkumulieren. Es wurde festgestellt, dass einige Vakuolen in Pflanzenzellen geringe Mengen an hydrolytischen Enzymen enthalten, und es wird vermutet, dass diese aus dem Golgi-Komplex stammen.
10. Pigmentbildung:
In vielen Tumor- und Krebszellen von Säugetieren wurde der Golgi-Komplex als Ursprungsort von Pigmentkörnchen (Melanin) beschrieben.
11. Regulierung des Flüssigkeitshaushaltes
Es wurde eine Homologie zwischen dem Golgi-Komplex und der kontraktilen Vakuole der unteren Metazoa und Protozoen vorgeschlagen. Die kontraktile Vakuole stößt überschüssiges Wasser aus der Zelle aus. In bestimmten Protozoen befasst sich der Golgi-Komplex auch mit der Regulierung des Flüssigkeitshaushalts.
Herkunft des Golgi-Komplexes:
Es wurden drei verschiedene Quellen vorgeschlagen, aus denen der neue Golgi-Komplex entstehen kann:
1. Vom endoplasmatischen Retikulum :
Essner und Novikøff (1962) und Beams and Kessel (1968) haben vorgeschlagen, dass die Golgi-Zisternen aus der Notaufnahme stammen. Das rauhe endoplasmatische Retikulum verliert nach der Synthese spezifischer Proteine Ribosomen und wandelt sich in glattes ER um. Kleine transitorische Vesikel klemmen sich vom glatten ER ab. Diese wandern zu Dictyosomen um.
Beim Erreichen des sich bildenden Gesichtes des Dictyosoms verschmelzen diese zu neuen Zisternen und tragen so zu ihrem Wachstum bei. Durch die Verschmelzung dieser Vesikel bilden sich ständig neue Zisternen auf der sich bildenden Fläche und auf der reifenden Fläche zerfallen die alten Zisternen in sekretorische Vesikel. So zeigt Golgi ein Phänomen der Membranströmung. '
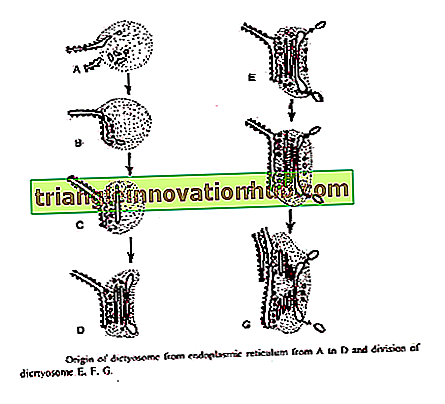

Bouch (1965) beschrieb den Ursprung von Golgi aus der äußeren Membran der Kernhülle in Braunalgen. Vesikel werden von der äußeren Kernmembran abgequetscht, die sich zu Zisternen auf der sich bildenden Fläche des Dictyosoms zusammenfügt.
Das Vorhandensein von Ausschlusszonen in Verbindung mit glatten ER oder Kernmembran, das Auftreten von Ausschlusszonen in ruhenden Samen höherer Pflanzen und die Bildung von Dictyosomen aus diesen Zonen in keimenden Samen liefern Beweise für die beiden oben genannten Theorien über den Ursprung von Dictyosom
3. Durch die Aufteilung von vorbestehendem Dictyosom :
Es wurde beobachtet, dass während der Zellteilung sowohl bei Pflanzen als auch bei Tieren die Anzahl der Dictyosomen zunimmt und die Anzahl der Dictyosomen in jeder Tochterzelle unmittelbar nach der Teilung nahezu gleich der Anzahl in der Stammzelle vor der Teilung ist, aus Dosen und anderen direkten Beobachtungen an den sich teilenden Zellen Es wurde vermutet, dass sich auch Diktyosomen während der Zellteilung teilen.